

Lactate produced by glycogenolysis in astrocytes regulates memory processing
- PMID: 22180782
- PMCID: PMC3236748
- DOI: 10.1371/journal.pone.0028427
Lactate produced by glycogenolysis in astrocytes regulates memory processing
Abstract
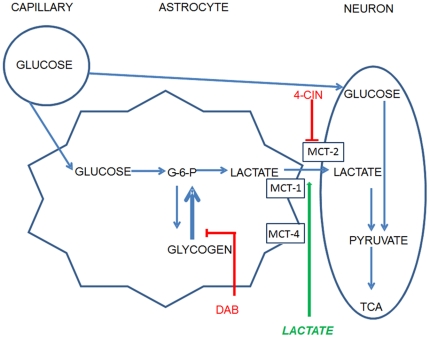
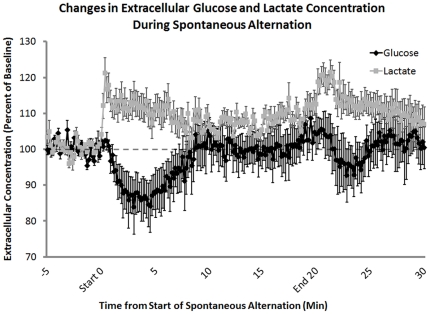
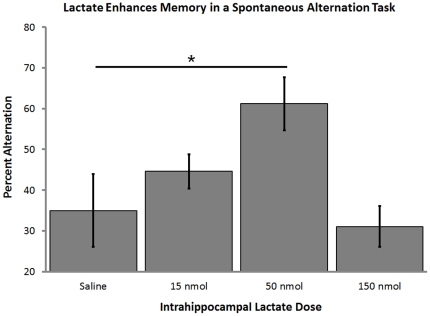
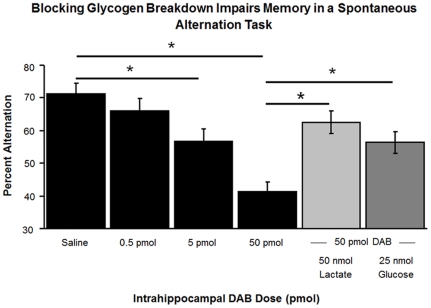
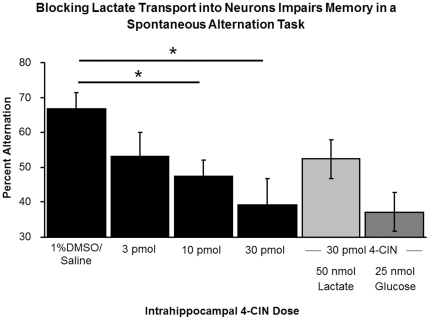
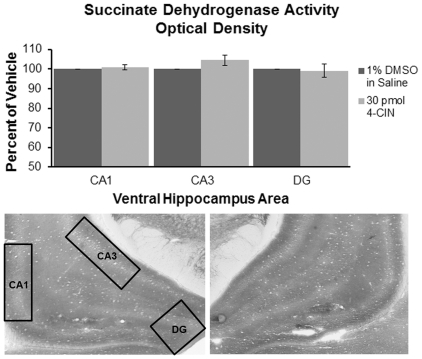
When administered either systemically or centrally, glucose is a potent enhancer of memory processes. Measures of glucose levels in extracellular fluid in the rat hippocampus during memory tests reveal that these levels are dynamic, decreasing in response to memory tasks and loads; exogenous glucose blocks these decreases and enhances memory. The present experiments test the hypothesis that glucose enhancement of memory is mediated by glycogen storage and then metabolism to lactate in astrocytes, which provide lactate to neurons as an energy substrate. Sensitive bioprobes were used to measure brain glucose and lactate levels in 1-sec samples. Extracellular glucose decreased and lactate increased while rats performed a spatial working memory task. Intrahippocampal infusions of lactate enhanced memory in this task. In addition, pharmacological inhibition of astrocytic glycogenolysis impaired memory and this impairment was reversed by administration of lactate or glucose, both of which can provide lactate to neurons in the absence of glycogenolysis. Pharmacological block of the monocarboxylate transporter responsible for lactate uptake into neurons also impaired memory and this impairment was not reversed by either glucose or lactate. These findings support the view that astrocytes regulate memory formation by controlling the provision of lactate to support neuronal functions.
Conflict of interest statement
Figures
References
-
- Messier C. Glucose improvement of memory: a review. European Journal of Pharmacology. 2004;490:33–57. - PubMed
-
- Gold PE. The many faces of amnesia. Learning & Memory. 2006;13:506–514. - PubMed
-
- Korol DL, Gold PE. Epinephrine converts long-term potentiation from transient to durable form in awake rats. Hippocampus. 2008;18:81–91. - PubMed
-
- Benton D, Owens D. Blood glucose and human memory. Psychopharmacology. 1993;113:83–88. - PubMed
-
- Benton D, Owens DS, Parker PY. Blood glucose influences memory and attention in young adults. Neuropsychologia. 1994;32:595–607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
