

Proteasomes and protein conjugation across domains of life
- PMID: 22183254
- PMCID: PMC3291102
- DOI: 10.1038/nrmicro2696
Proteasomes and protein conjugation across domains of life
Abstract
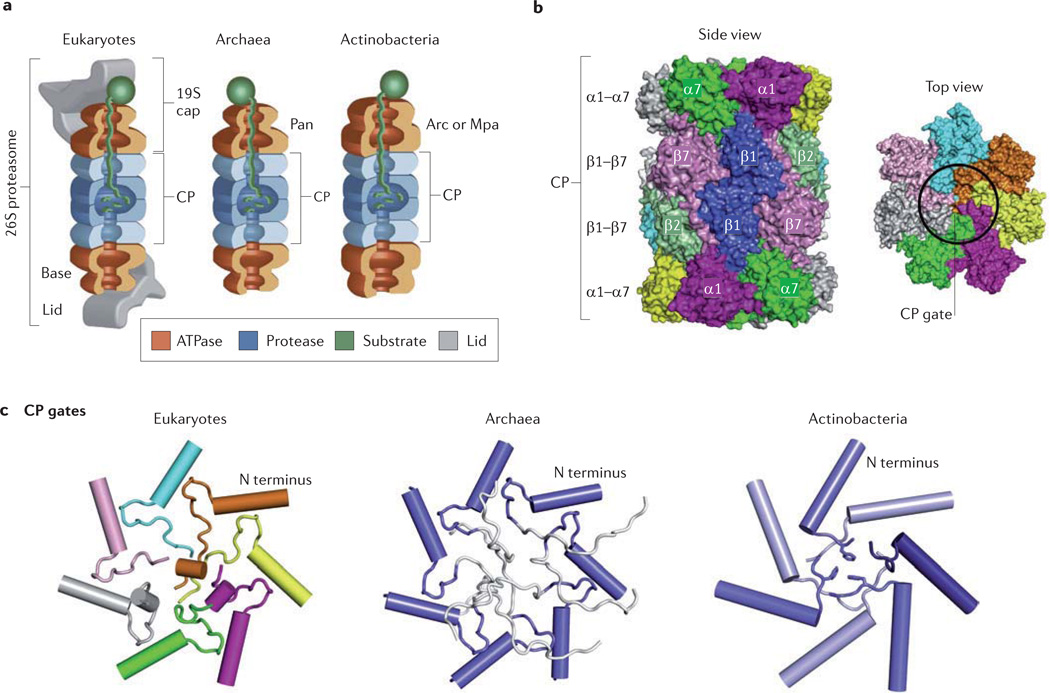
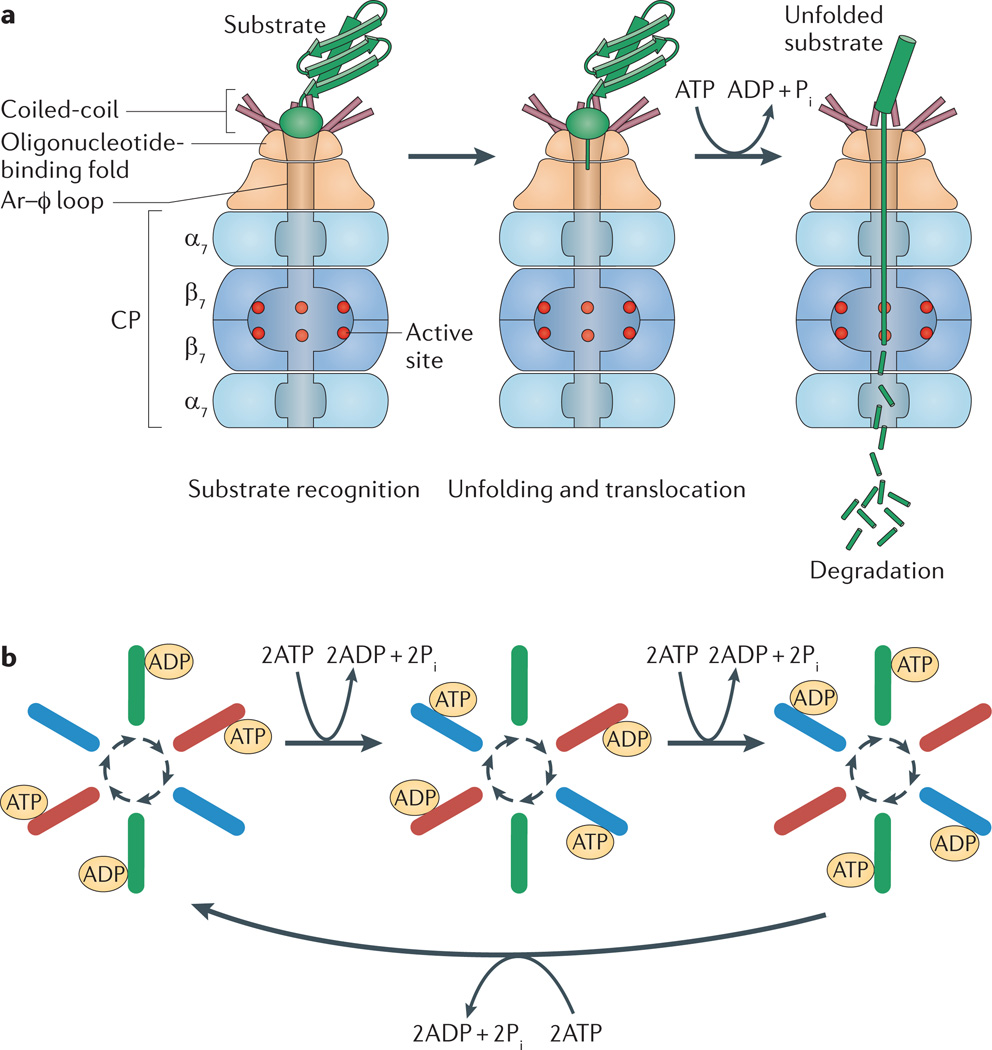
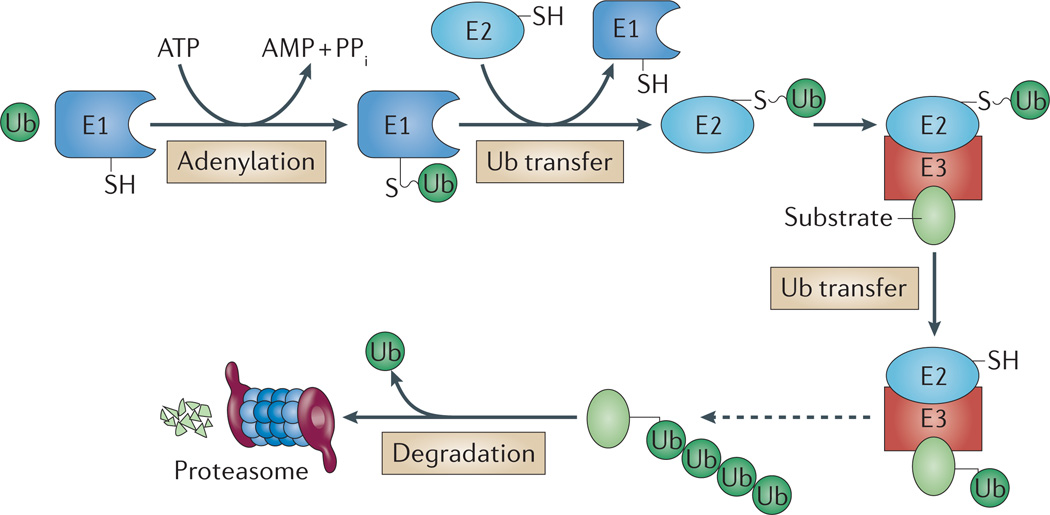
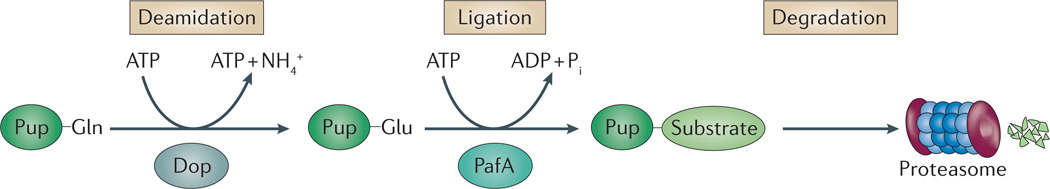
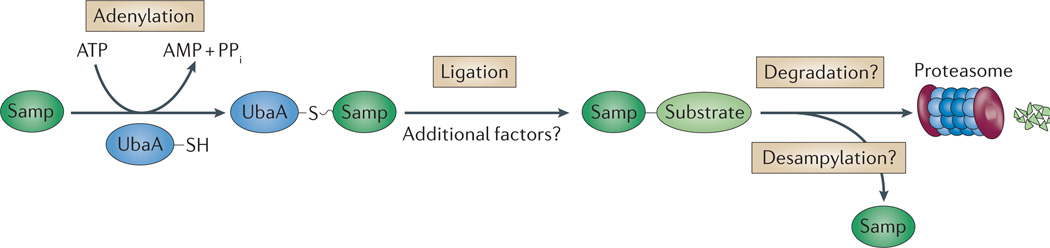
Like other energy-dependent proteases, proteasomes, which are found across the three domains of life, are self-compartmentalized and important in the early steps of proteolysis. Proteasomes degrade improperly synthesized, damaged or misfolded proteins and hydrolyse regulatory proteins that must be specifically removed or cleaved for cell signalling. In eukaryotes, proteins are typically targeted for proteasome-mediated destruction through polyubiquitylation, although ubiquitin-independent pathways also exist. Interestingly, actinobacteria and archaea also covalently attach small proteins (prokaryotic ubiquitin-like protein (Pup) and small archaeal modifier proteins (Samps), respectively) to certain proteins, and this may serve to target the modified proteins for degradation by proteasomes.
Figures
References
-
- Volker C, Lupas AN. Molecular evolution of proteasomes. Curr. Top. Microbiol. Immunol. 2002;268:1–22. - PubMed
-
- Mehnert M, Sommer T, Jarosch E. ERAD ubiquitin ligases: multifunctional tools for protein quality control and waste disposal in the endoplasmic reticulum. Bioessays. 2010;32:905–913. - PubMed
-
- Barford D. Structure, function and mechanism of the anaphase promoting complex (APC/C) Q. Rev. Biophys. 2011;44:153–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
