

Ca(v)2 channels mediate low and high voltage-activated calcium currents in Drosophila motoneurons
- PMID: 22183725
- PMCID: PMC3381312
- DOI: 10.1113/jphysiol.2011.222836
Ca(v)2 channels mediate low and high voltage-activated calcium currents in Drosophila motoneurons
Abstract
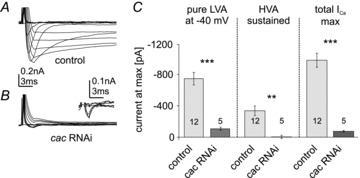
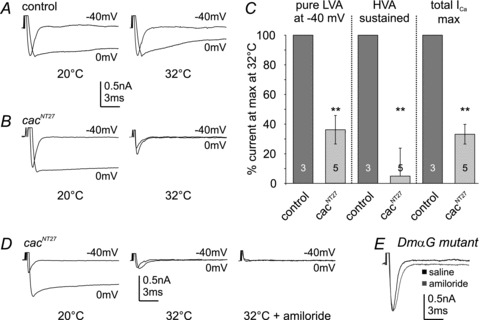
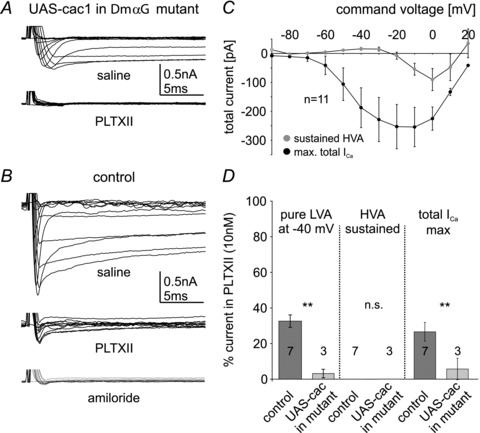
Different blends of membrane currents underlie distinct functions of neurons in the brain. A major step towards understanding neuronal function, therefore, is to identify the genes that encode different ionic currents. This study combined in situ patch clamp recordings of somatodendritic calcium currents in an identified adult Drosophila motoneuron with targeted genetic manipulation. Voltage clamp recordings revealed transient low voltage-activated (LVA) currents with activation between –60 mV and –70 mV as well as high voltage-activated (HVA) current with an activation voltage around –30 mV. LVA could be fully inactivated by prepulses to –50 mV and was partially amiloride sensitive. Recordings from newly generated mutant flies demonstrated that DmαG (Ca(v)3 homolog) encoded the amiloride-sensitive portion of the transient LVA calcium current. We further demonstrated that the Ca(v)2 homolog, Dmca1A, mediated the amiloride-insensitive component of LVA current. This novel role of Ca(v)2 channels was substantiated by patch clamp recordings from conditional mutants, RNAi knock-downs, and following Dmca1A overexpression. In addition, we show that Dmca1A underlies the HVA somatodendritic calcium currents in vivo. Therefore, the Drosophila Ca(v)2 homolog, Dmca1A, underlies HVA and LVA somatodendritic calcium currents in the same neuron. Interestingly, DmαG is required for regulating LVA and HVA derived from Dmca1A in vivo. In summary, each vertebrate gene family for voltage-gated calcium channels is represented by a single gene in Drosophila, namely Dmca1D (Ca(v)1), Dmca1A (Ca(v)2) and DmαG (Ca(v)3), but the commonly held view that LVA calcium currents are usually mediated by Ca(v)3 rather than Ca(v)2 channels may require reconsideration.
Figures
Similar articles
-
Characterization of voltage-dependent Ca2+ currents in identified Drosophila motoneurons in situ.J Neurophysiol. 2008 Aug;100(2):868-78. doi: 10.1152/jn.90464.2008. Epub 2008 Jun 11. J Neurophysiol. 2008. PMID: 18550721 Free PMC article.
-
Calcium conductances and their role in the firing behavior of neonatal rat hypoglossal motoneurons.J Neurophysiol. 1993 Jun;69(6):2137-49. doi: 10.1152/jn.1993.69.6.2137. J Neurophysiol. 1993. PMID: 8394413
-
Whole-cell and single-channel calcium currents in guinea pig basal forebrain neurons.J Neurophysiol. 1994 Jun;71(6):2359-76. doi: 10.1152/jn.1994.71.6.2359. J Neurophysiol. 1994. PMID: 7931521
-
Voltage-dependent potentiation of low-voltage-activated Ca2+ channel currents in cultured rat bone marrow cells.J Physiol. 1995 Dec 15;489 ( Pt 3)(Pt 3):649-61. doi: 10.1113/jphysiol.1995.sp021080. J Physiol. 1995. PMID: 8788931 Free PMC article.
-
Voltage-dependent calcium channels.Gen Physiol Biophys. 2005 Jun;24 Suppl 1:1-78. Gen Physiol Biophys. 2005. PMID: 16096350 Review.
Cited by
-
Neuronal calcium spikes enable vector inversion in the Drosophila brain.bioRxiv [Preprint]. 2023 Nov 28:2023.11.24.568537. doi: 10.1101/2023.11.24.568537. bioRxiv. 2023. PMID: 38077032 Free PMC article. Preprint.
-
Different functions of two putative Drosophila α2δ subunits in the same identified motoneurons.Sci Rep. 2020 Aug 13;10(1):13670. doi: 10.1038/s41598-020-69748-8. Sci Rep. 2020. PMID: 32792569 Free PMC article.
-
Putative excitatory and putative inhibitory inputs are localised in different dendritic domains in a Drosophila flight motoneuron.Eur J Neurosci. 2013 Mar;37(6):860-75. doi: 10.1111/ejn.12104. Epub 2012 Dec 27. Eur J Neurosci. 2013. PMID: 23279094 Free PMC article.
-
In Vivo Calcium Signaling during Synaptic Refinement at the Drosophila Neuromuscular Junction.J Neurosci. 2017 May 31;37(22):5511-5526. doi: 10.1523/JNEUROSCI.2922-16.2017. Epub 2017 May 5. J Neurosci. 2017. PMID: 28476946 Free PMC article.
-
Restoration of Motor Defects Caused by Loss of Drosophila TDP-43 by Expression of the Voltage-Gated Calcium Channel, Cacophony, in Central Neurons.J Neurosci. 2017 Sep 27;37(39):9486-9497. doi: 10.1523/JNEUROSCI.0554-17.2017. Epub 2017 Aug 28. J Neurosci. 2017. PMID: 28847811 Free PMC article.
References
-
- Baines RA, Pym EC. Determinants of electrical properties in developing neurons. Semin Cell Dev Biol. 2006;17:12–19. - PubMed
-
- Bleakman D, Bowman D, Bath CP, Brust PF, Johnson EC, Deal CR, Miller RJ, Ellis SB, Harpold MM, Hans M, Grantham CJ. Characteristics of a human N-type calcium channel expressed in HEK293 cells. Neuropharmacology. 1995;34:753–765. - PubMed
-
- Bock G, Gebhart M, Scharinger A, Jangsangthong W, Busquet P, Poggiani C, S Sartori, Mangoni ME, Sinnegger-Brauns MJ, Herzig S, Striessnig J, Koschak A. Functional properties of a newly identified C-terminal splice variant of Cav1.3 L-type Ca2+ channels. J Biol Chem. 2011;286:42736–42748. - PMC - PubMed
-
- Boerner J, Duch C. Average shape standard atlas for the adult Drosophila ventral nerve cord. J Comp Neurol. 2010;518:2437–2455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases