

Desmosomal cadherins utilize distinct kinesins for assembly into desmosomes
- PMID: 22184201
- PMCID: PMC3246898
- DOI: 10.1083/jcb.201106057
Desmosomal cadherins utilize distinct kinesins for assembly into desmosomes
Erratum in
- J Cell Biol. 2012 Jan 23;196(2):297
- J Cell Biol. 2013 Jun 24;201(7):1085
Abstract
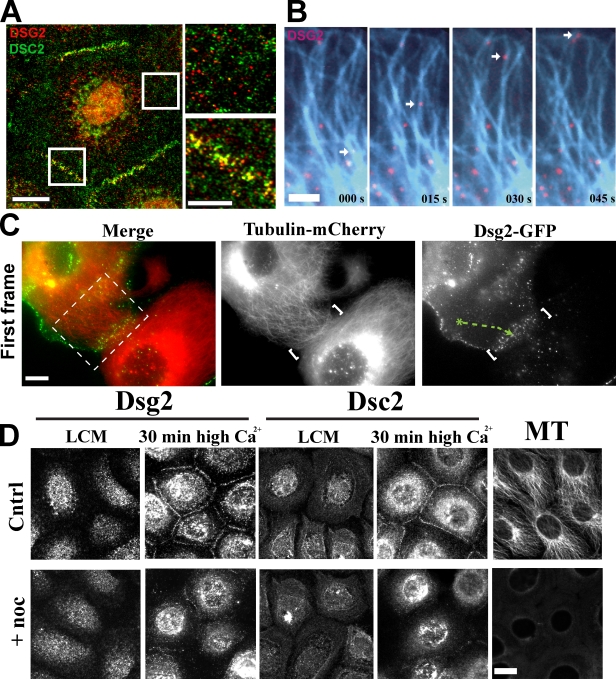
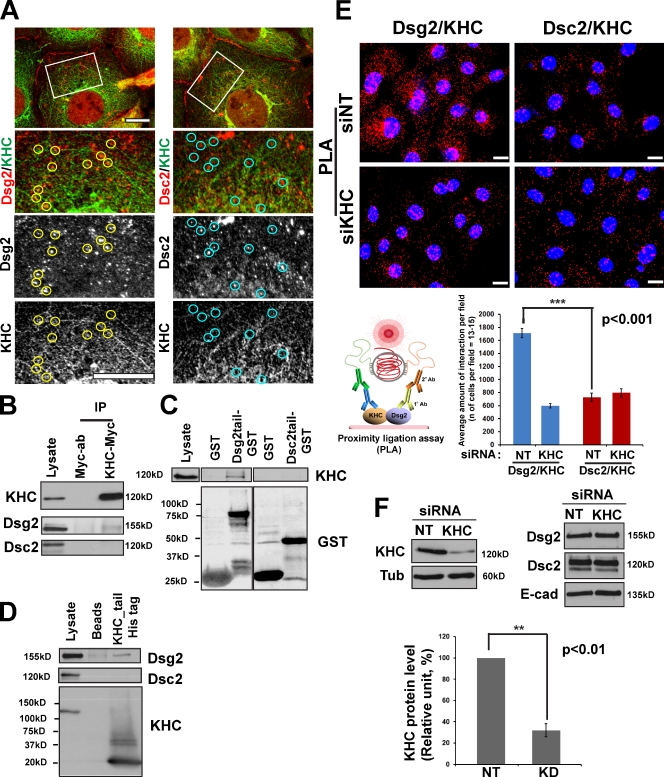
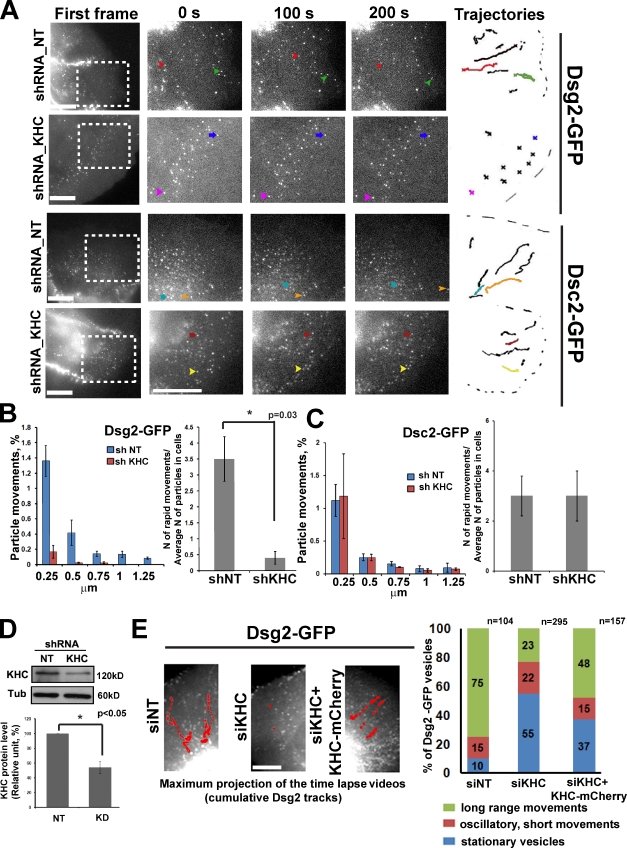
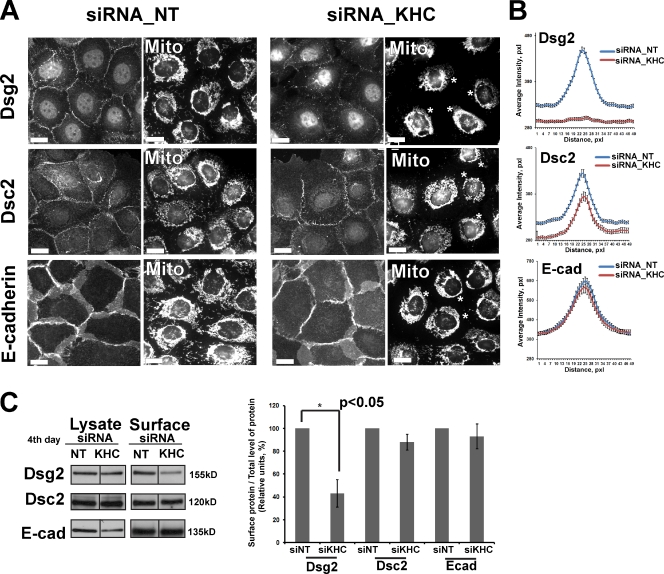
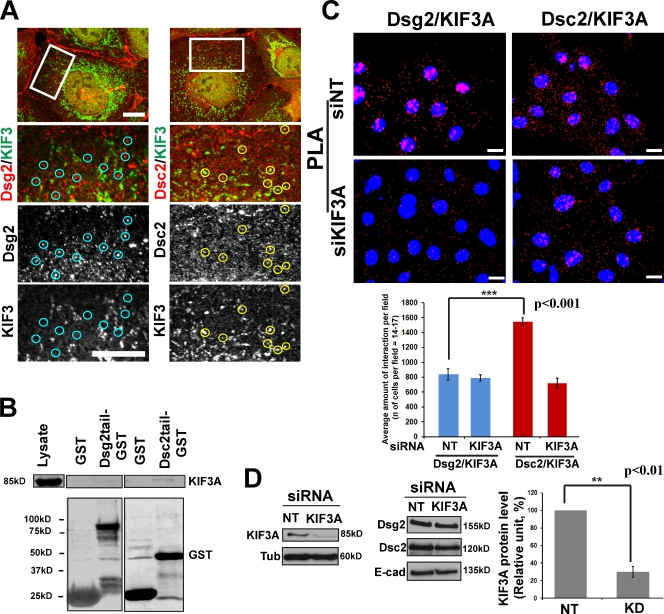
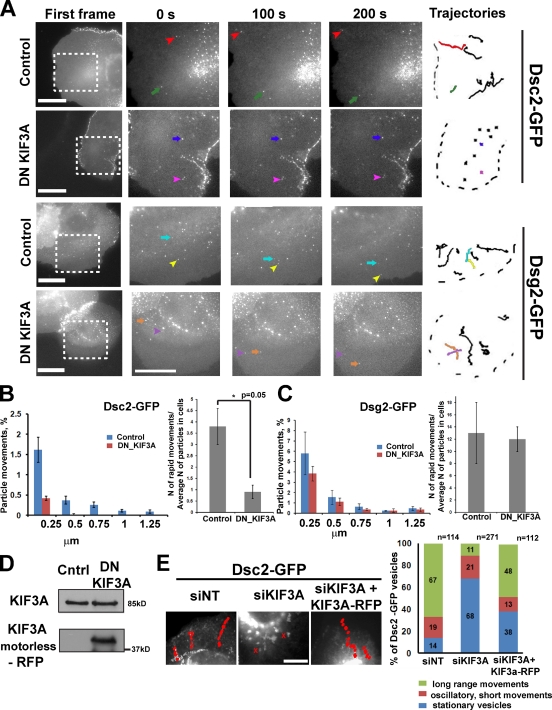
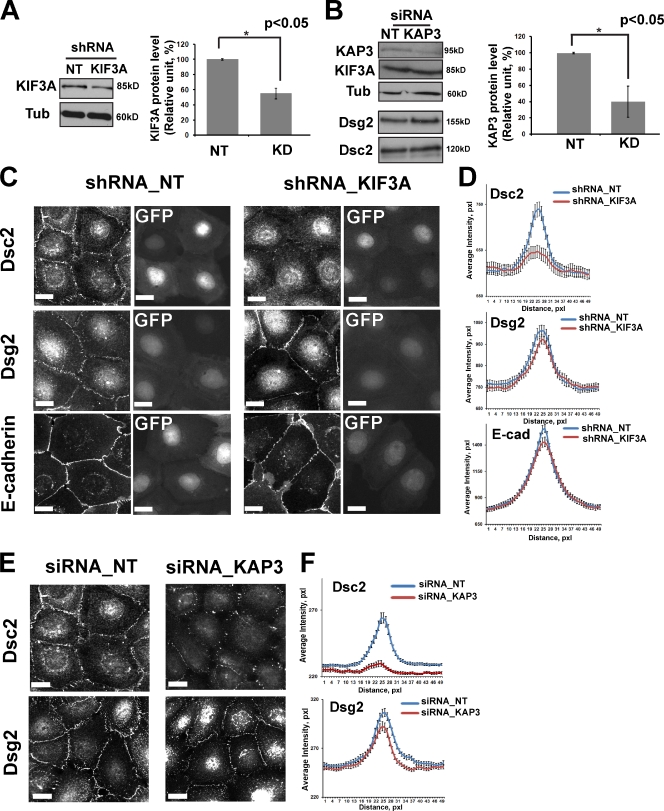
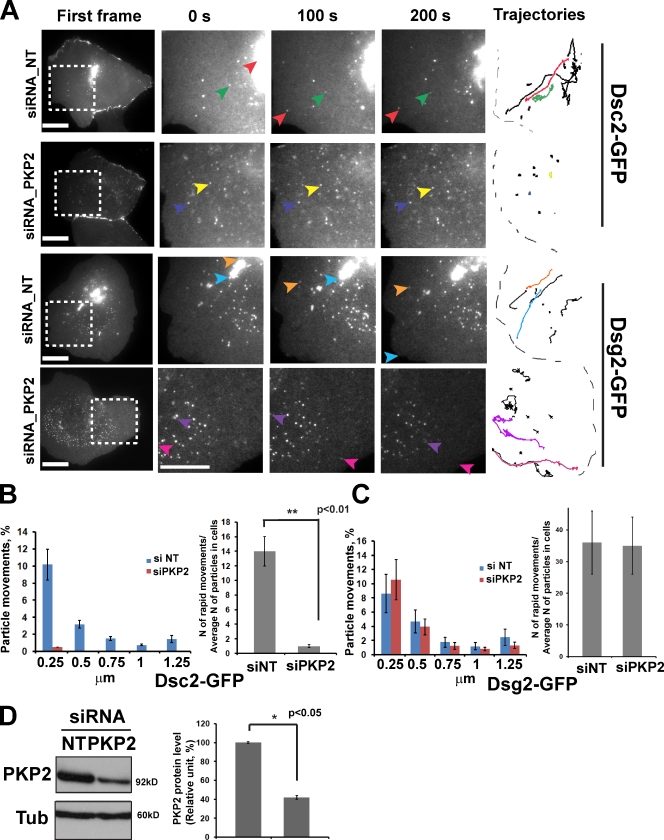
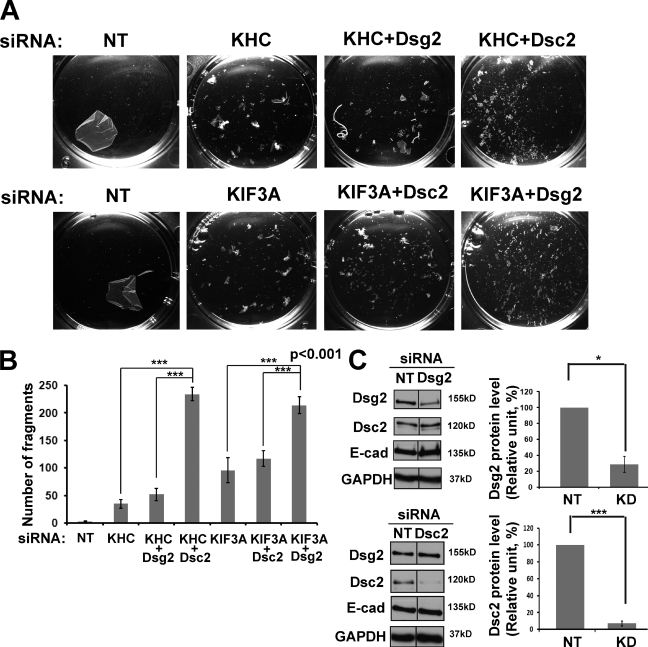
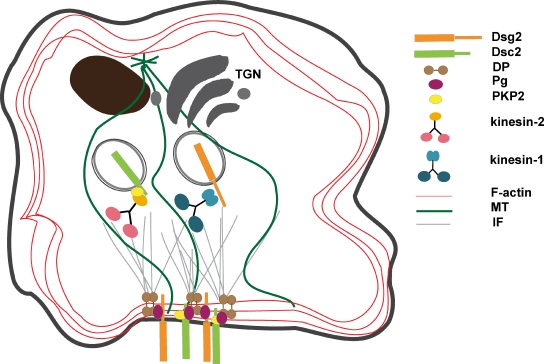
The desmosomal cadherins, desmogleins (Dsgs) and desmocollins (Dscs), comprise the adhesive core of intercellular junctions known as desmosomes. Although these adhesion molecules are known to be critical for tissue integrity, mechanisms that coordinate their trafficking into intercellular junctions to regulate their proper ratio and distribution are unknown. We demonstrate that Dsg2 and Dsc2 both exhibit microtubule-dependent transport in epithelial cells but use distinct motors to traffic to the plasma membrane. Functional interference with kinesin-1 blocked Dsg2 transport, resulting in the assembly of Dsg2-deficient junctions with minimal impact on distribution of Dsc2 or desmosomal plaque components. In contrast, inhibiting kinesin-2 prevented Dsc2 movement and decreased its plasma membrane accumulation without affecting Dsg2 trafficking. Either kinesin-1 or -2 deficiency weakened intercellular adhesion, despite the maintenance of adherens junctions and other desmosome components at the plasma membrane. Differential regulation of desmosomal cadherin transport could provide a mechanism to tailor adhesion strength during tissue morphogenesis and remodeling.
Figures
References
-
- Angst B.D., Nilles L.A., Green K.J. 1990. Desmoplakin II expression is not restricted to stratified epithelia. J. Cell Sci. 97:247–257 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
