

Gene order and chromosome dynamics coordinate spatiotemporal gene expression during the bacterial growth cycle
- PMID: 22184251
- PMCID: PMC3258614
- DOI: 10.1073/pnas.1108229109
Gene order and chromosome dynamics coordinate spatiotemporal gene expression during the bacterial growth cycle
Abstract
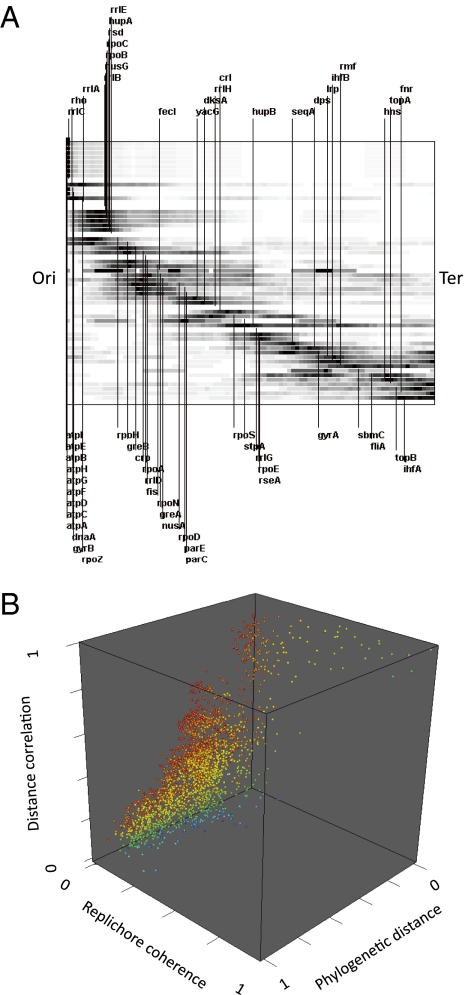
In Escherichia coli crosstalk between DNA supercoiling, nucleoid-associated proteins and major RNA polymerase σ initiation factors regulates growth phase-dependent gene transcription. We show that the highly conserved spatial ordering of relevant genes along the chromosomal replichores largely corresponds both to their temporal expression patterns during growth and to an inferred gradient of DNA superhelical density from the origin to the terminus. Genes implicated in similar functions are related mainly in trans across the chromosomal replichores, whereas DNA-binding transcriptional regulators interact predominantly with targets in cis along the replichores. We also demonstrate that macrodomains (the individual structural partitions of the chromosome) are regulated differently. We infer that spatial and temporal variation of DNA superhelicity during the growth cycle coordinates oxygen and nutrient availability with global chromosome structure, thus providing a mechanistic insight into how the organization of a complete bacterial chromosome encodes a spatiotemporal program integrating DNA replication and global gene expression.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
DNA thermodynamic stability and supercoil dynamics determine the gene expression program during the bacterial growth cycle.Mol Biosyst. 2013 Jul;9(7):1643-51. doi: 10.1039/c3mb25515h. Epub 2013 Mar 14. Mol Biosyst. 2013. PMID: 23493878
-
Spatiotemporal Coupling of DNA Supercoiling and Genomic Sequence Organization-A Timing Chain for the Bacterial Growth Cycle?Biomolecules. 2022 Jun 15;12(6):831. doi: 10.3390/biom12060831. Biomolecules. 2022. PMID: 35740956 Free PMC article. Review.
-
Analog regulation of metabolic demand.BMC Syst Biol. 2011 Mar 15;5:40. doi: 10.1186/1752-0509-5-40. BMC Syst Biol. 2011. PMID: 21406074 Free PMC article.
-
Genome scale patterns of supercoiling in a bacterial chromosome.Nat Commun. 2016 Mar 30;7:11055. doi: 10.1038/ncomms11055. Nat Commun. 2016. PMID: 27025941 Free PMC article.
-
Partitioning of the Escherichia coli chromosome: superhelicity and condensation.Biochimie. 2001 Jan;83(1):41-8. doi: 10.1016/s0300-9084(00)01204-9. Biochimie. 2001. PMID: 11254973 Review.
Cited by
-
The nucleoid as a smart polymer.Front Microbiol. 2015 May 8;6:424. doi: 10.3389/fmicb.2015.00424. eCollection 2015. Front Microbiol. 2015. PMID: 26005440 Free PMC article. No abstract available.
-
The highly conserved chromosomal periodicity of transcriptomes and the correlation of its amplitude with the growth rate in Escherichia coli.DNA Res. 2020 Jun 1;27(3):dsaa018. doi: 10.1093/dnares/dsaa018. DNA Res. 2020. PMID: 32866232 Free PMC article.
-
Comparative Genomics of Interreplichore Translocations in Bacteria: A Measure of Chromosome Topology?G3 (Bethesda). 2016 Jun 1;6(6):1597-606. doi: 10.1534/g3.116.028274. G3 (Bethesda). 2016. PMID: 27172194 Free PMC article.
-
Unveiling DNA structural features of promoters associated with various types of TSSs in prokaryotic transcriptomes and their role in gene expression.DNA Res. 2017 Feb 1;24(1):25-35. doi: 10.1093/dnares/dsw045. DNA Res. 2017. PMID: 27803028 Free PMC article.
-
Variable DNA topology is an epigenetic generator of physiological heterogeneity in bacterial populations.Mol Microbiol. 2023 Jan;119(1):19-28. doi: 10.1111/mmi.15014. Epub 2022 Dec 30. Mol Microbiol. 2023. PMID: 36565252 Free PMC article. Review.
References
-
- Hsieh LS, Burger RM, Drlica K. Bacterial DNA supercoiling and [ATP]/[ADP]. Changes associated with a transition to anaerobic growth. J Mol Biol. 1991;219:443–450. - PubMed
-
- Bordes P, et al. DNA supercoiling contributes to disconnect sigmaS accumulation from sigmaS-dependent transcription in Escherichia coli. Mol Microbiol. 2003;48:561–571. - PubMed
-
- Rimsky S, Travers A. Pervasive regulation of nucleoid structure and function by nucleoid-associated proteins. Curr Opin Microbiol. 2011;14:136–141. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
