

Review
doi: 10.1242/dev.067074.
Shaping sound in space: the regulation of inner ear patterning
Affiliations
- PMID: 22186725
- PMCID: PMC3243092
- DOI: 10.1242/dev.067074
Item in Clipboard
Review
Shaping sound in space: the regulation of inner ear patterning
Development.
2012 Jan.
Erratum in
- Development. 2012 Feb;139(4):826
Abstract
The inner ear is one of the most morphologically elaborate tissues in vertebrates, containing a group of mechanosensitive sensory organs that mediate hearing and balance. These organs are arranged precisely in space and contain intricately patterned sensory epithelia. Here, we review recent studies of inner ear development and patterning which reveal that multiple stages of ear development - ranging from its early induction from the embryonic ectoderm to the establishment of the three cardinal axes and the fine-grained arrangement of sensory cells - are orchestrated by gradients of signaling molecules.
Figures
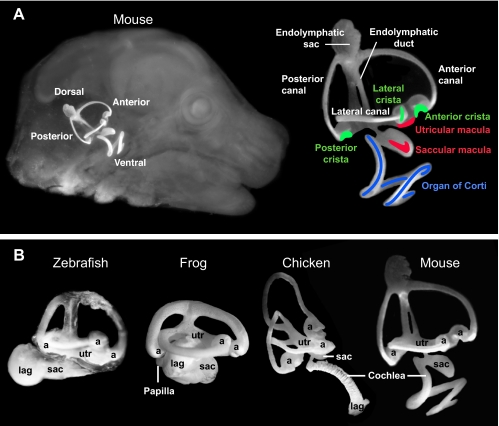
Inner ear anatomy. The interconnected fluid cavities of the inner ear can be revealed following injection of white paint in fixed and cleared specimens. (A) The mouse ear viewed in situ (left) and in detail (right) at E15.5. The approximate locations of sensory patches are indicated: organ of Corti, blue; maculae, red; cristae, green. (B) Lateral views of zebrafish, frog, chicken and mouse inner ears with the major chambers labeled. The orthogonal placement of semicircular canals dorsally is a shared feature of inner ears, although basal vertebrates such as lampreys have two, rather than the full complement of three, canals (not shown). At the end of each canal is an enlarged space, the ampulla (a). The endolymphatic duct projects dorsally and enlarges into a sac (shown only for mouse). Vestibular macular sensory organs are located in the utricle (utr), saccule (sac) and lagena (lag), although in fish the saccular macula has been co-opted to sense sound. In all species, the ventral ear houses a hearing organ, with frequency sensitivity that varies systematically along its length. In mammals, hearing is subserved by the organ of Corti located in the coiled cochlea. In archosaurs, lizards and amphibians, elongated sensory organs called papillae have evolved to sense hearing.
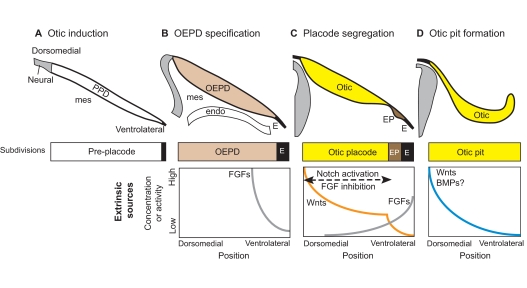
Otic induction from the pre-placode. (A) Otic induction requires extrinsic sources of secreted molecules originating from neural tissue, mesenchyme (mes) and pharyngeal endoderm (endo). (B) FGFs act on the pre-placodal domain (PPD) to specify the otic-epibranchial placode domain (OEPD) as separate from ectoderm (E). (C) The OEPD field is further segregated into the otic placode (Otic) and the epibranchial placodes (EP). The otic placode forms under the influence of high Wnts and low FGF signaling. This begins with a Wnt gradient that develops a sharp transition point through feedback loops involving Notch activation and FGF inhibition. (D) By the time the otic pit begins to invaginate, the dorsomedial domain may already be receiving higher Wnt and BMP signals from the adjacent hindbrain, thereby initiating both dorsal-ventral (DV) and mediolateral patterning (not shown). Note that in this and subsequent figures, illustrations of gradients of signaling proteins or their resulting activities are not based on direct observation but rather are speculative, taking into consideration results from experimental embryology, known gene expression patterns and/or phenotypes resulting from perturbations of signaling pathways using drugs or in mutant mice.
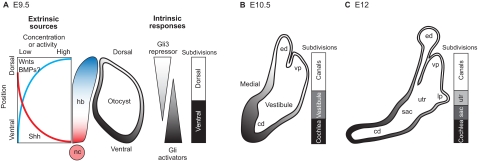
Dorsoventral axial patterning during inner ear development. (A) At E9.5, the mouse otocyst is positioned to receive secreted signals that originate asymmetrically from the adjacent hindbrain (hb) and notochord (nc). During otocyst development, gradients of sonic hedgehog (Shh) target genes are observed along the DV axis (gray shading). A working model is shown in which the otic epithelium experiences opposing gradients of Gli activators (high ventrally) and the Gli3 repressor (high dorsally) to establish positional information across the entire DV axis. Wnt signaling is also a major contributor to the patterning of dorsal inner ear structures. BMPs are also present but have not yet been shown to have a role in DV axis specification of the inner ear. (B,C) Between E10.5 and E12, the mouse otocyst enlarges and becomes progressively segregated along the DV axis into chambers and canals. Transcription factor expression domains mark dorsal versus ventral territories at E9.5-10.5. However, prior to overt morphogenesis, the lack of markers for a transitional domain (the vestibule) or for the saccular (sac) and utricular (utr) chambers, means that the diagrammatic representation of the sequential subdivision of these regions is speculative. Note that the 3-fold expansion in the size of the inner ear during these stages is not illustrated. cd, cochlear duct; ed, endolymphatic duct; lp, lateral canal plate; vp, vertical canal plate.
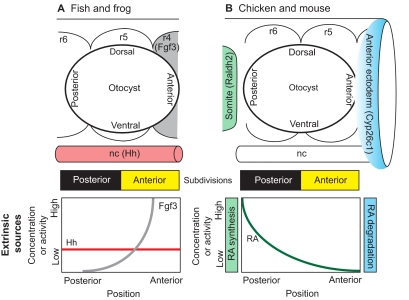
Anterior-posterior axial patterning in the inner ear. Extrinsic sources of signaling molecules that pattern the otic placode/vesicle differ between anamniotes and amniotes. (A) Zebrafish and Xenopus otic vesicles begin with a mirror-image symmetric prepattern of otic tissue that is centered along the anterior-posterior (AP) axis adjacent to rhombomere (r) 5. The symmetric template is then independently endowed with posterior identity via hedgehog (Hh) signaling originating from the notochord (nc) and anterior identity via Fgf3 signaling originating from rhombomere 4. (B) In chicken and mouse, the otocyst is centered next to the r5/r6 boundary. AP positional information is influenced by an extrinsic gradient of retinoic acid (RA). This gradient arises because of asymmetry in the synthesis and degradation of RA by enzymes (Raldh2 and Cyp26c1) present in the somatic mesoderm and anterior ectoderm, respectively. There are no lineage data to suggest that the otic field becomes fully subdivided into anterior and posterior halves, as shown schematically by the yellow and black bars for both fish/frog and chicken/mouse. However, the observation of mirror-image symmetry along this axis in mutants or following experimental manipulations supports the idea that there are two distinct compartments.
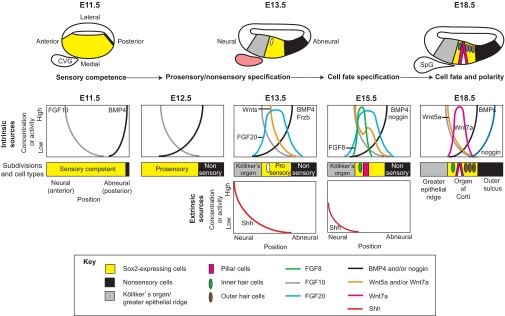
Cochlear patterning. The medial wall of the mouse cochlea undergoes sequential subdivisions under the influence of morphogen family members. Between E11.5 and E13.5, Sox2-expressing sensory-competent cells (yellow) become confined to a central domain, called the prosensory domain, and cease cell division. By E18.5, this region differentiates into the organ of Corti, which is composed of a single row of inner hair cells (green), a pair of pillar cells (pink) and three rows of outer hair cells (brown) that lie above Sox2-expressing supporting cells (yellow). The organ of Corti is flanked by two nonsensory regions: Kölliker's organ (gray) on the neural side and the outer sulcus (black) on the abneural side. Genes for several morphogens and signaling molecules, including FGFs, Wnts and BMPs, are expressed in discrete territories within the epithelium (intrinsic sources), from which they could diffuse to form gradients to mediate positional information in adjacent cells. Other signals, such as Shh, arise from nearby tissues (extrinsic sources). The cochleovestibular ganglion (CVG), for example, is an extrinsic source of Shh, which is presumed to diffuse into the epithelium to repress sensory differentiation of Kölliker's organ. The CVG will separate into a vestibular ganglion (not shown) and the spiral ganglion (SpG) of the cochlea. Shh expression is transient and is not present in the SpG on E18.5. Predicted gradients for these intrinsic and extrinsic signals are shown for the mid-base, reflecting a longitudinal location about one-quarter of the distance to the tip of the cochlea.
References
-
- Abello G., Khatri S., Radosevic M., Scotting P. J., Giraldez F., Alsina B. (2010). Independent regulation of Sox3 and Lmx1b by FGF and BMP signaling influences the neurogenic and non-neurogenic domains in the chick otic placode. Dev. Biol. 339, 166-178 - PubMed
-
- Baker C. V., Bronner-Fraser M. (2001). Vertebrate cranial placodes I. Embryonic induction. Dev. Biol. 232, 1-61 - PubMed
-
- Bertrand N., Dahmane N. (2006). Sonic hedgehog signaling in forebrain development and its interactions with pathways that modify its effects. Trends Cell Biol. 16, 597-605 - PubMed
-
- Bok J., Bronner-Fraser M., Wu D. K. (2005). Role of the hindbrain in dorsoventral but not anteroposterior axial specification of the inner ear. Development 132, 2115-2124 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
