

Competition for FcRn-mediated transport gives rise to short half-life of human IgG3 and offers therapeutic potential
- PMID: 22186895
- PMCID: PMC3247843
- DOI: 10.1038/ncomms1608
Competition for FcRn-mediated transport gives rise to short half-life of human IgG3 and offers therapeutic potential
Abstract
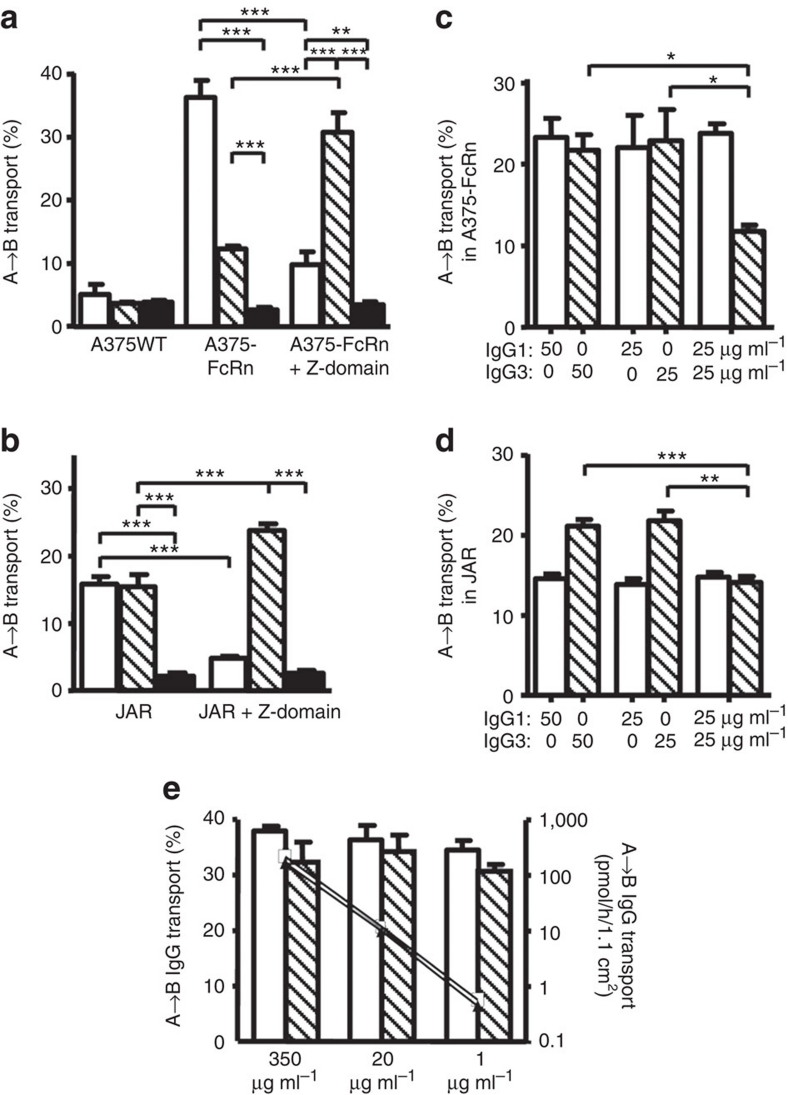
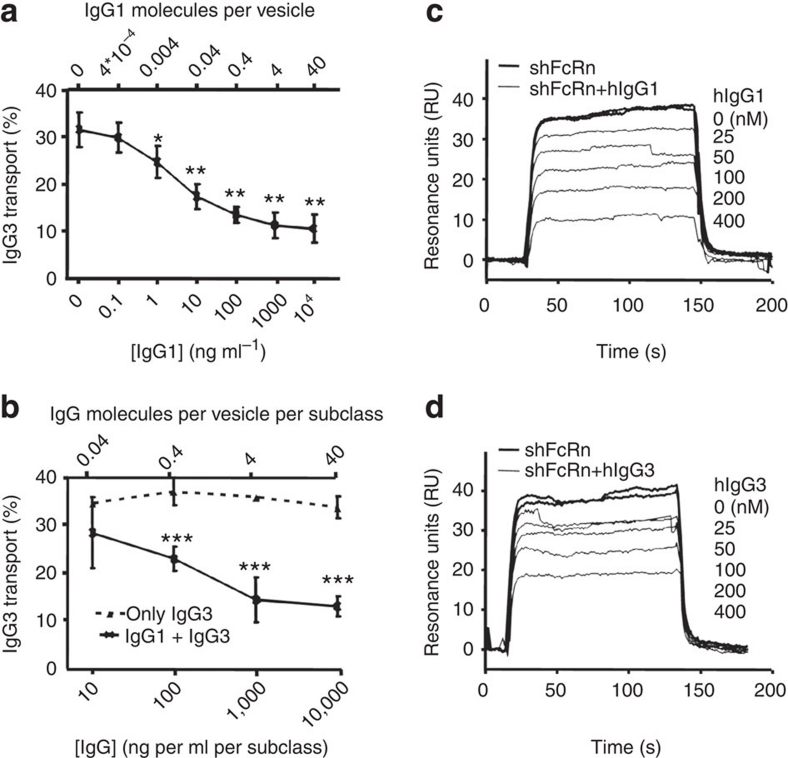
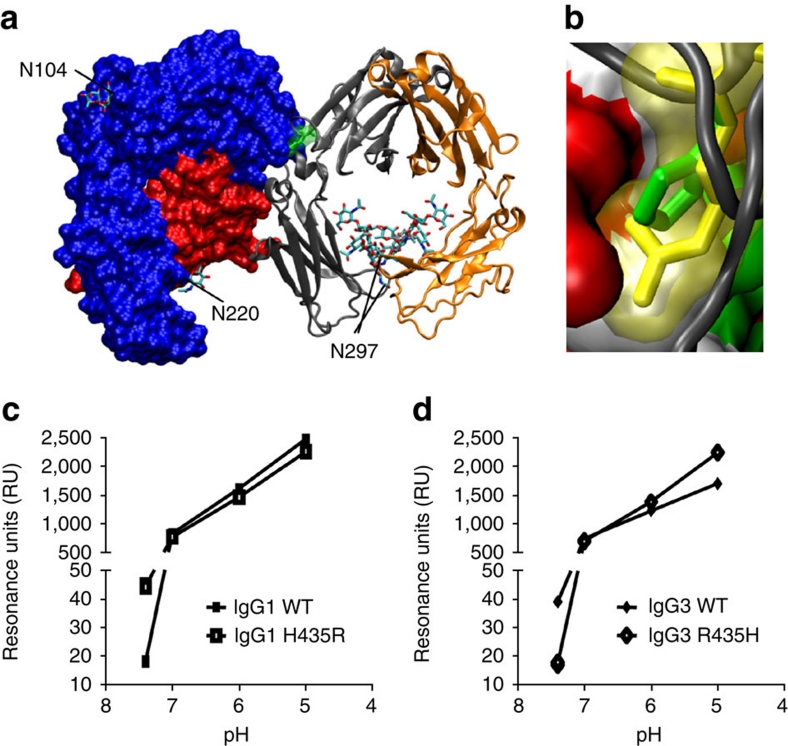
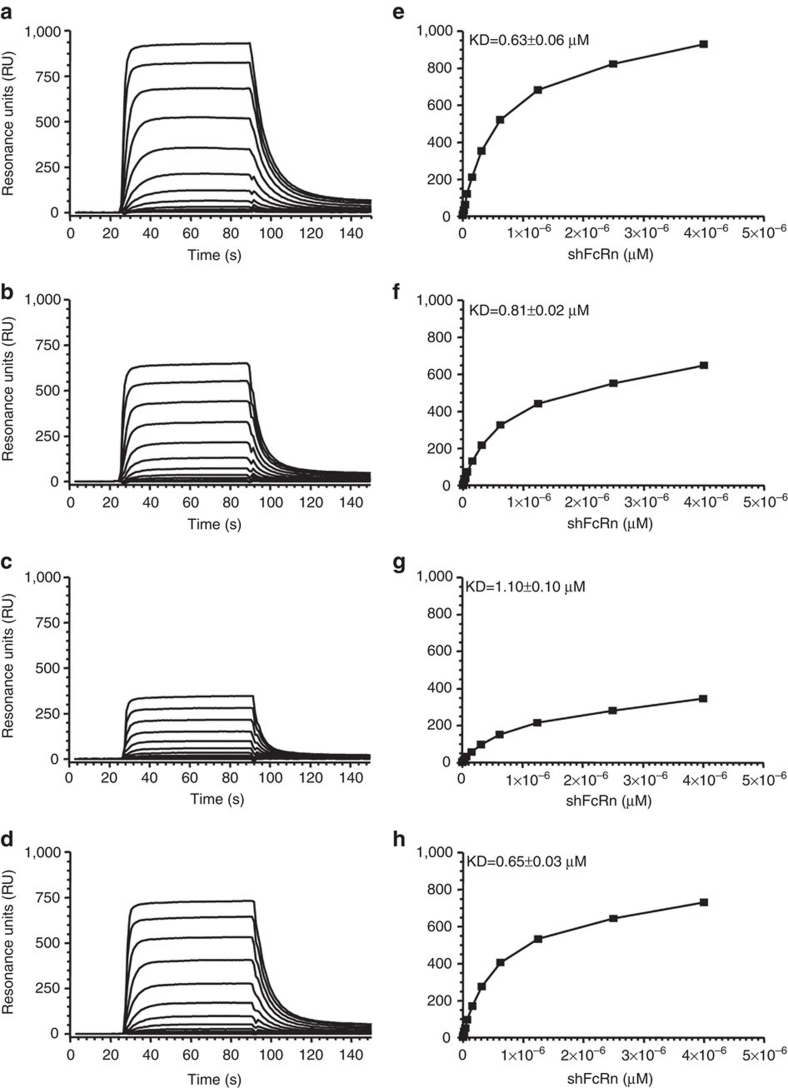
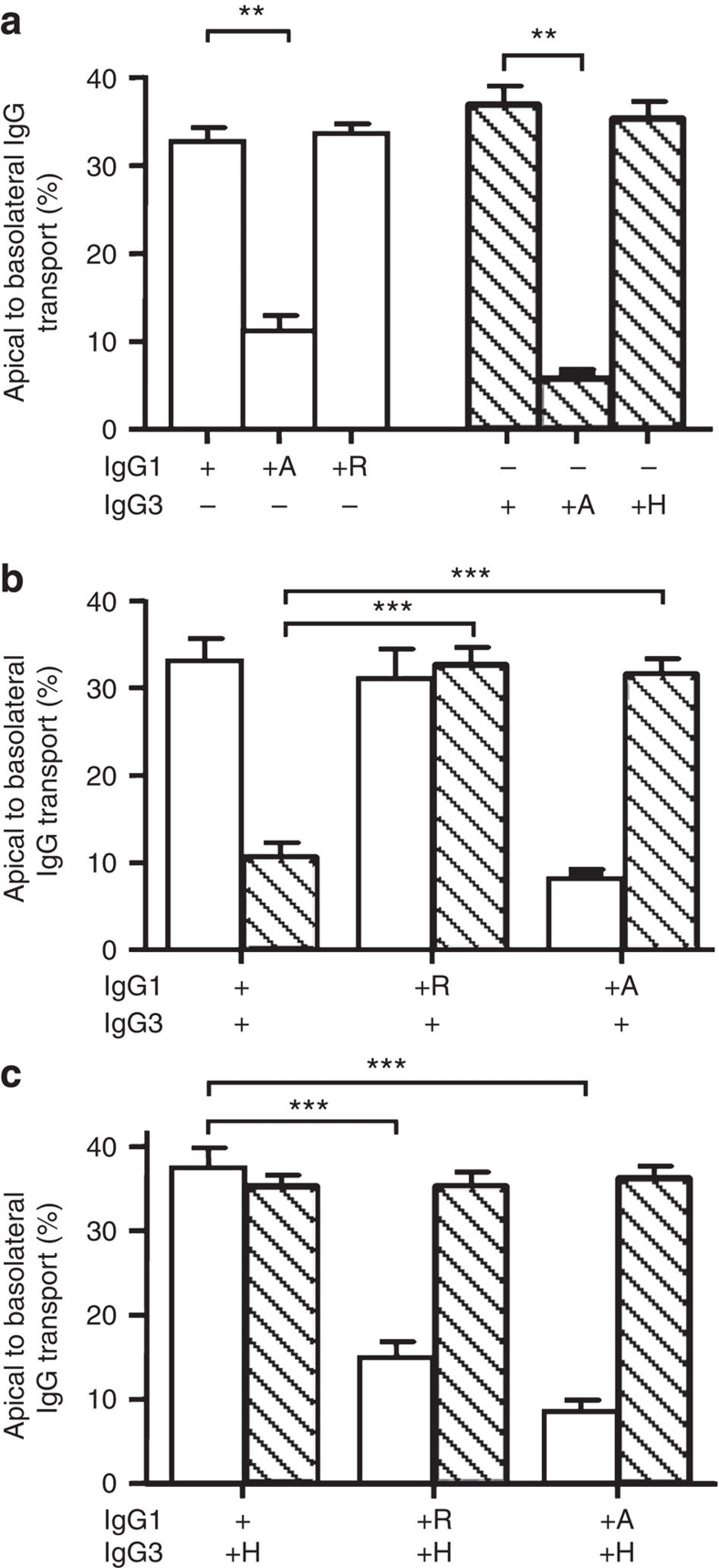
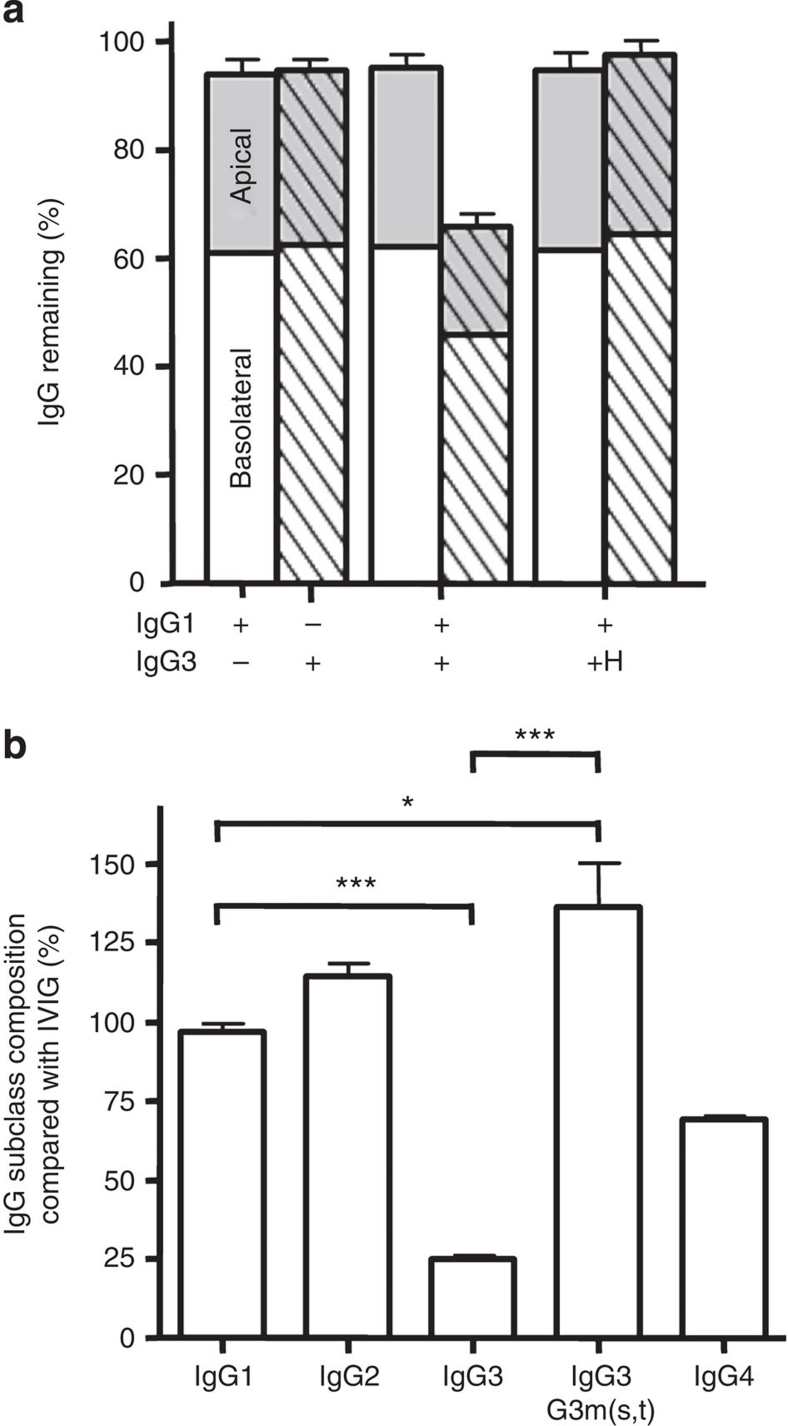
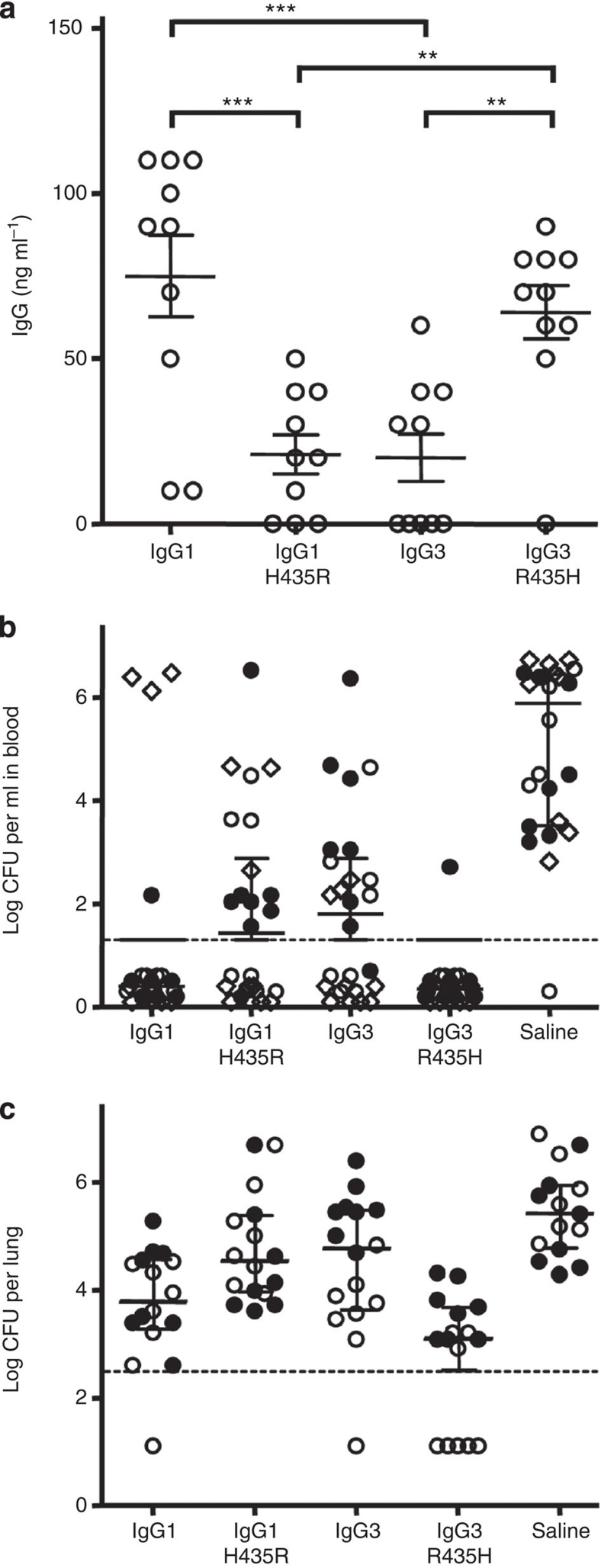
Human IgG3 displays the strongest effector functions of all IgG subclasses but has a short half-life for unresolved reasons. Here we show that IgG3 binds to IgG-salvage receptor (FcRn), but that FcRn-mediated transport and rescue of IgG3 is inhibited in the presence of IgG1 due to intracellular competition between IgG1 and IgG3. We reveal that this occurs because of a single amino acid difference at position 435, where IgG3 has an arginine instead of the histidine found in all other IgG subclasses. While the presence of R435 in IgG increases binding to FcRn at neutral pH, it decreases binding at acidic pH, affecting the rescue efficiency-but only in the presence of H435-IgG. Importantly, we show that in humans the half-life of the H435-containing IgG3 allotype is comparable to IgG1. H435-IgG3 also gave enhanced protection against a pneumococcal challenge in mice, demonstrating H435-IgG3 to be a candidate for monoclonal antibody therapies.
Figures
References
-
- Bruhns P. et al. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood 113, 3716–3725 (2009). - PubMed
-
- Salfeld J. G. Isotype selection in antibody engineering. Nat. Biotechnol. 25, 1369–1372 (2007). - PubMed
-
- Natsume A. et al. Engineered antibodies of IgG1/IgG3 mixed isotype with enhanced cytotoxic activities. Cancer Res. 68, 3863–3872 (2008). - PubMed
-
- Jefferis R. Antibody therapeutics: isotype and glycoform selection. Expert Opin. Biol. Ther. 7, 1401–1413 (2007). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
