

The food additive vanillic acid controls transgene expression in mammalian cells and mice
- PMID: 22187155
- PMCID: PMC3300003
- DOI: 10.1093/nar/gkr1251
The food additive vanillic acid controls transgene expression in mammalian cells and mice
Abstract
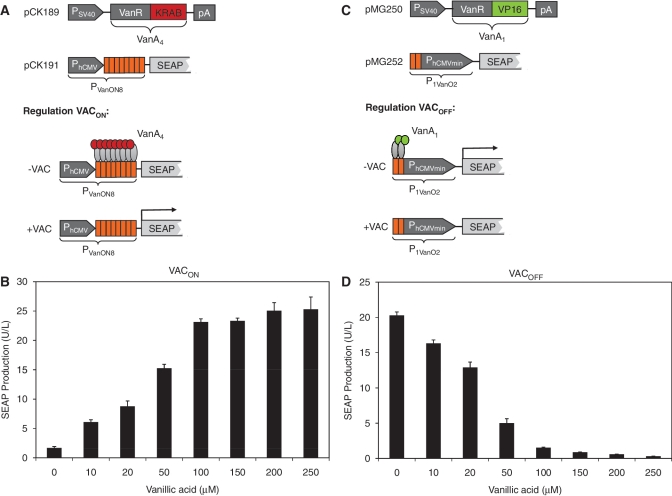
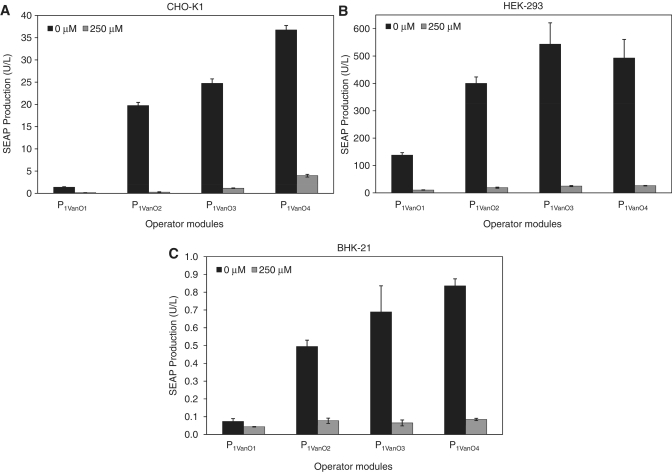
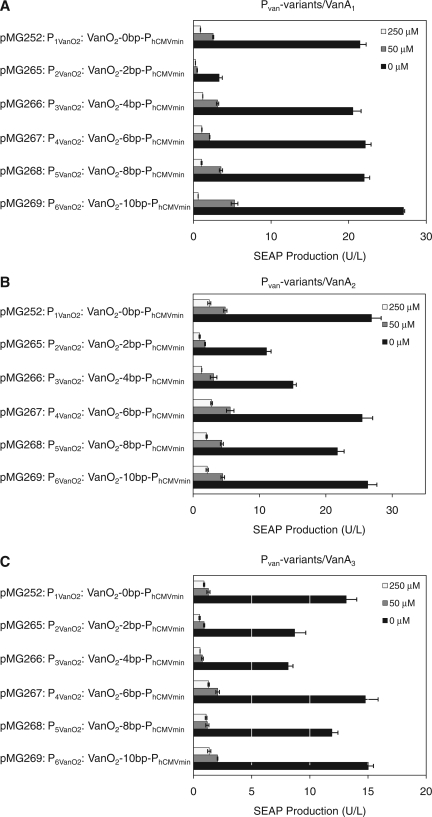
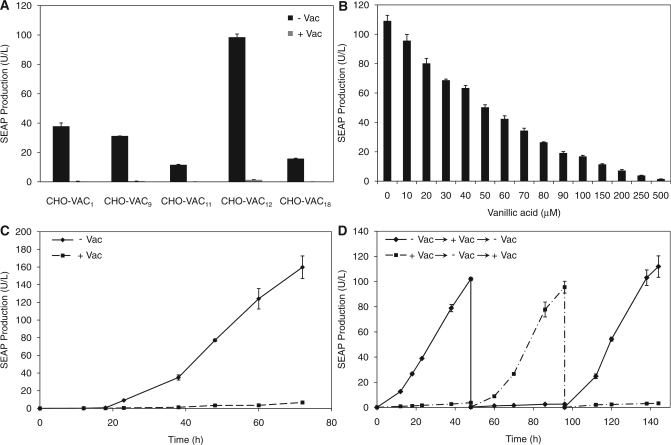
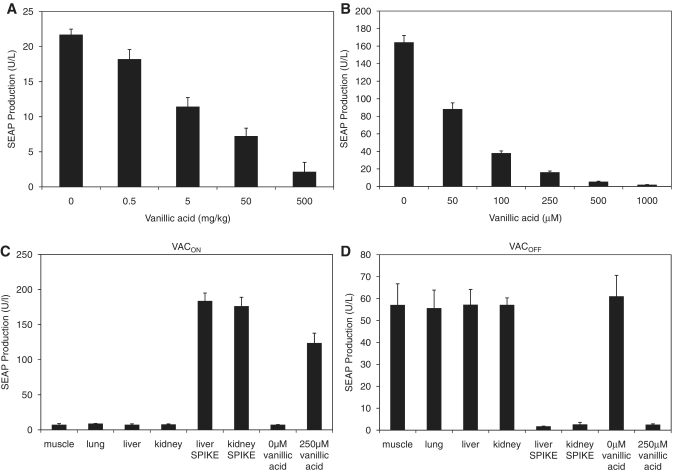
Trigger-inducible transcription-control devices that reversibly fine-tune transgene expression in response to molecular cues have significantly advanced the rational reprogramming of mammalian cells. When designed for use in future gene- and cell-based therapies the trigger molecules have to be carefully chosen in order to provide maximum specificity, minimal side-effects and optimal pharmacokinetics in a mammalian organism. Capitalizing on control components that enable Caulobacter crescentus to metabolize vanillic acid originating from lignin degradation that occurs in its oligotrophic freshwater habitat, we have designed synthetic devices that specifically adjust transgene expression in mammalian cells when exposed to vanillic acid. Even in mice transgene expression was robust, precise and tunable in response to vanillic acid. As a licensed food additive that is regularly consumed by humans via flavoured convenience food and specific fresh vegetable and fruits, vanillic acid can be considered as a safe trigger molecule that could be used for diet-controlled transgene expression in future gene- and cell-based therapies.
Figures
References
-
- Baumgartel K, Genoux D, Welzl H, Tweedie-Cullen RY, Koshibu K, Livingstone-Zatchej M, Mamie C, Mansuy IM. Control of the establishment of aversive memory by calcineurin and Zif268. Nat. Neurosci. 2008;11:572–578. - PubMed
-
- Gitzinger M, Parsons J, Reski R, Fussenegger M. Functional cross-kingdom conservation of mammalian and moss (Physcomitrella patens) transcription, translation and secretion machineries. Plant Biotechnol. J. 2009;7:73–86. - PubMed
-
- Ulmer JB, Valley U, Rappuoli R. Vaccine manufacturing: challenges and solutions. Nat. Biotechnol. 2006;24:1377–1383. - PubMed
-
- Weber W, Fussenegger M. Inducible product gene expression technology tailored to bioprocess engineering. Curr. Opin. Biotechnol. 2007;18:399–410. - PubMed
-
- Sharpless NE, Depinho RA. The mighty mouse: genetically engineered mouse models in cancer drug development. Nat. Rev. Drug Discov. 2006;5:741–754. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
