

Probing transcription-specific outputs of β-catenin in vivo
- PMID: 22190459
- PMCID: PMC3248684
- DOI: 10.1101/gad.181289.111
Probing transcription-specific outputs of β-catenin in vivo
Abstract
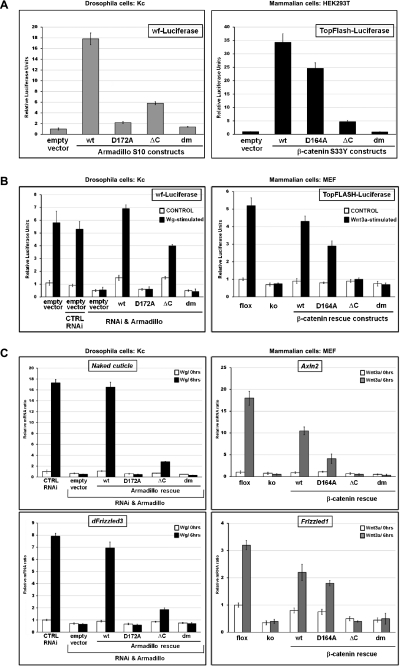
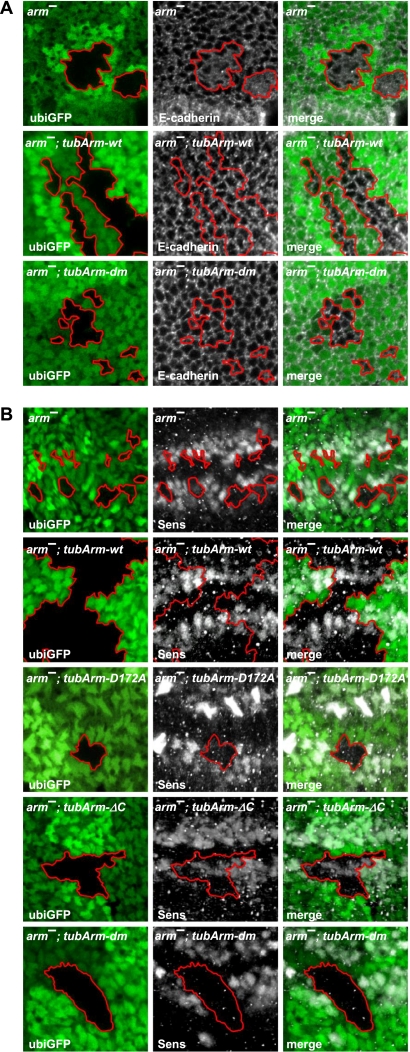
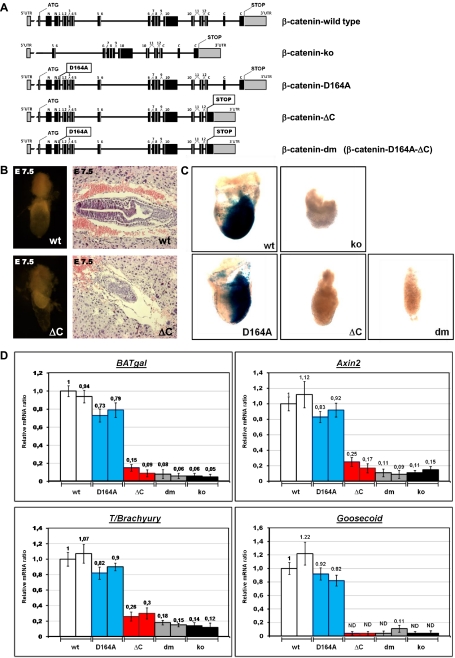
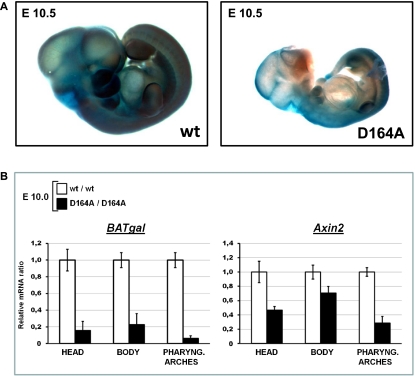
β-Catenin, apart from playing a cell-adhesive role, is a key nuclear effector of Wnt signaling. Based on activity assays in Drosophila, we generated mouse strains where the endogenous β-catenin protein is replaced by mutant forms, which retain the cell adhesion function but lack either or both of the N- and the C-terminal transcriptional outputs. The C-terminal activity is essential for mesoderm formation and proper gastrulation, whereas N-terminal outputs are required later during embryonic development. By combining the double-mutant β-catenin with a conditional null allele and a Wnt1-Cre driver, we probed the role of Wnt/β-catenin signaling in dorsal neural tube development. While loss of β-catenin protein in the neural tube results in severe cell adhesion defects, the morphology of cells and tissues expressing the double-mutant form is normal. Surprisingly, Wnt/β-catenin signaling activity only moderately regulates cell proliferation, but is crucial for maintaining neural progenitor identity and for neuronal differentiation in the dorsal spinal cord. Our model animals thus allow dissecting signaling and structural functions of β-catenin in vivo and provide the first genetic tool to generate cells and tissues that entirely and exclusively lack canonical Wnt pathway activity.
© 2011 by Cold Spring Harbor Laboratory Press
Figures
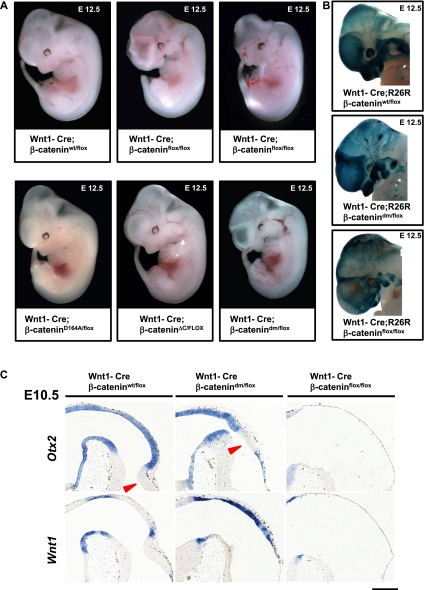
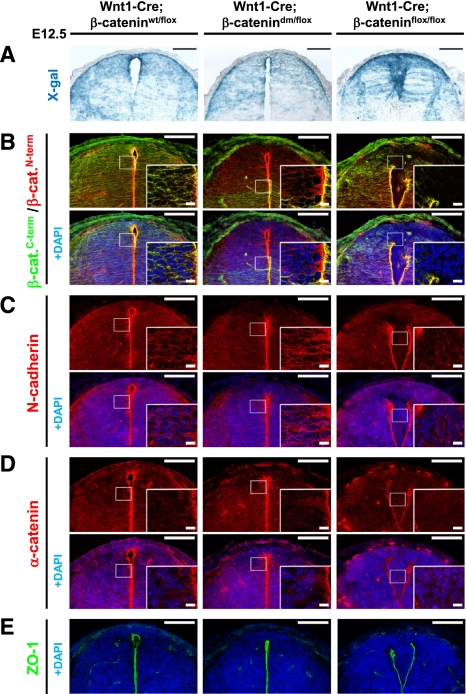
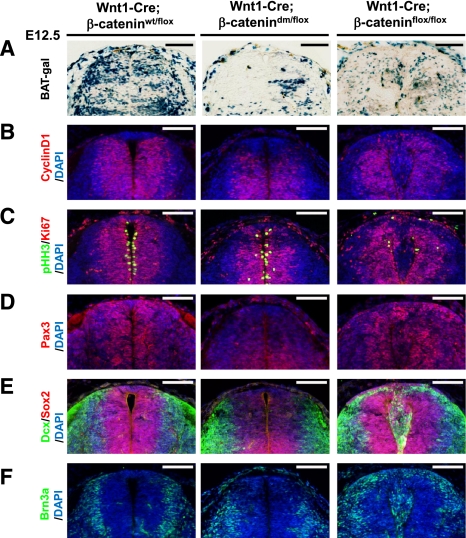
Comment in
-
Wnt signaling: the β-cat(enin)'s meow.Genes Dev. 2012 Jan 15;26(2):105-9. doi: 10.1101/gad.185280.111. Genes Dev. 2012. PMID: 22279043 Free PMC article.
References
-
- Arnold SJ, Robertson EJ 2009. Making a commitment: Cell lineage allocation and axis patterning in the early mouse embryo. Nat Rev Mol Cell Biol 10: 91–103 - PubMed
-
- Arnold SJ, Stappert J, Bauer A, Kispert A, Herrmann BG, Kemler R 2000. Brachyury is a target gene of the Wnt/β-catenin signaling pathway. Mech Dev 91: 249–258 - PubMed
-
- Basler K, Struhl G 1994. Compartment boundaries and the control of Drosophila limb pattern by hedgehog protein. Nature 368: 208–214 - PubMed
-
- Brault V, Moore R, Kutsch S, Ishibashi M, Rowitch DH, McMahon AP, Sommer L, Boussadia O, Kemler R 2001. Inactivation of the β-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development 128: 1253–1264 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases