

Conformational properties of cardiolipin-bound cytochrome c
- PMID: 22190488
- PMCID: PMC3252944
- DOI: 10.1073/pnas.1112312108
Conformational properties of cardiolipin-bound cytochrome c
Abstract
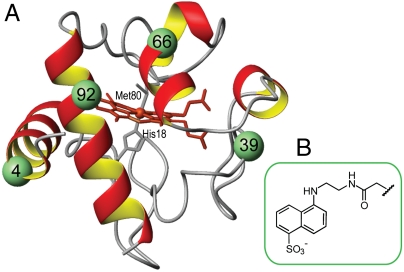
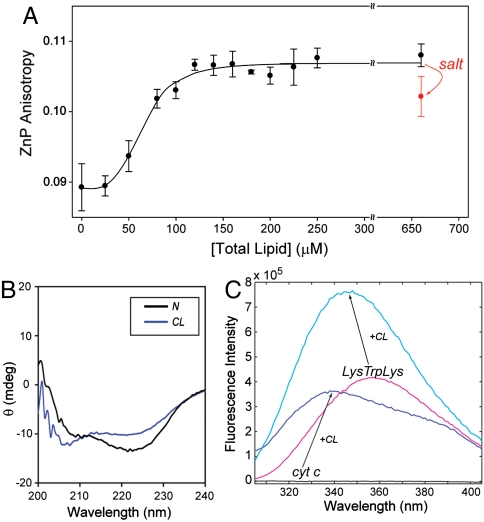
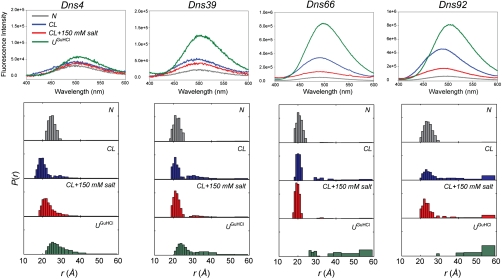
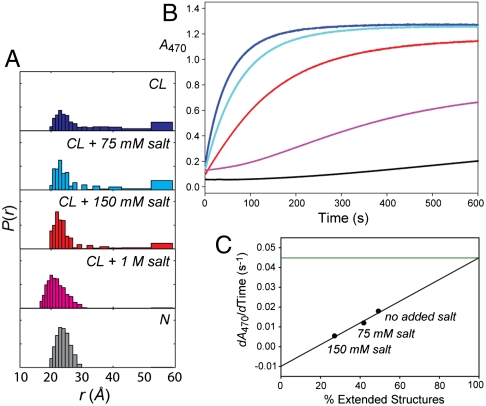
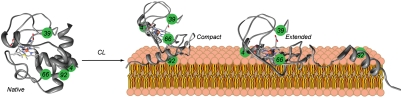
Interactions of cytochrome c (cyt c) with cardiolipin (CL) are important for both electron transfer and apoptotic functions of this protein. A sluggish peroxidase in its native state, when bound to CL, cyt c catalyzes CL peroxidation, which contributes to the protein apoptotic release. The heterogeneous CL-bound cyt c ensemble is difficult to characterize with traditional structural methods and ensemble-averaged probes. We have employed time-resolved FRET measurements to evaluate structural properties of the CL-bound protein in four dansyl (Dns)-labeled variants of horse heart cyt c. The Dns decay curves and extracted Dns-to-heme distance distributions P(r) reveal a conformational diversity of the CL-bound cyt c ensemble with distinct populations of the polypeptide structures that vary in their degree of protein unfolding. A fraction of the ensemble is substantially unfolded, with Dns-to-heme distances resembling those in the guanidine hydrochloride-denatured state. These largely open cyt c structures likely dominate the peroxidase activity of the CL-bound cyt c ensemble. Site variations in P(r) distributions uncover structural features of the CL-bound cyt c, rationalize previous findings, and implicate the prime role of electrostatic interactions, particularly with the protein C terminus, in the CL-induced unfolding.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ow YP, Green DR, Hao Z, Mak TW. Cytochrome c: functions beyond respiration. Nat Rev Mol Cell Biol. 2008;9:532–542. - PubMed
-
- Kagan VE, et al. Cytochrome c acts as a cardiolipin oxygenase required for release of proapoptotic factors. Nat Chem Biol. 2005;1:223–232. - PubMed
-
- Iverson SL, Orrenius S. The cardiolipin-cytochrome c interaction and the mitochondrial regulation of apoptosis. Arch Biochem Biophys. 2004;423:37–46. - PubMed
-
- Sinibaldi F, et al. Insights into cytochrome c-cardiolipin interaction. Role played by ionic strength. Biochemistry. 2008;47:6928–6935. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
