

"Antelope": a hybrid-logic model checker for branching-time Boolean GRN analysis
- PMID: 22192526
- PMCID: PMC3316443
- DOI: 10.1186/1471-2105-12-490
"Antelope": a hybrid-logic model checker for branching-time Boolean GRN analysis
Abstract
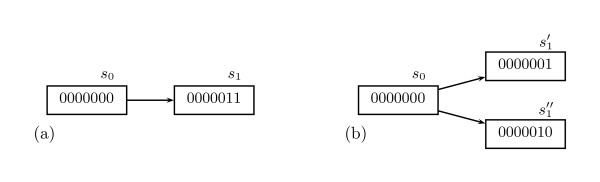
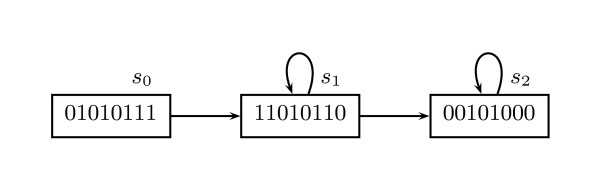
Background: In Thomas' formalism for modeling gene regulatory networks (GRNs), branching time, where a state can have more than one possible future, plays a prominent role. By representing a certain degree of unpredictability, branching time can model several important phenomena, such as (a) asynchrony, (b) incompletely specified behavior, and (c) interaction with the environment. Introducing more than one possible future for a state, however, creates a difficulty for ordinary simulators, because infinitely many paths may appear, limiting ordinary simulators to statistical conclusions. Model checkers for branching time, by contrast, are able to prove properties in the presence of infinitely many paths.
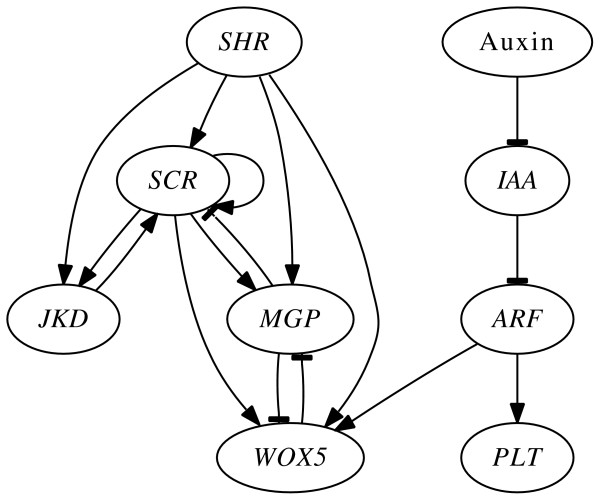
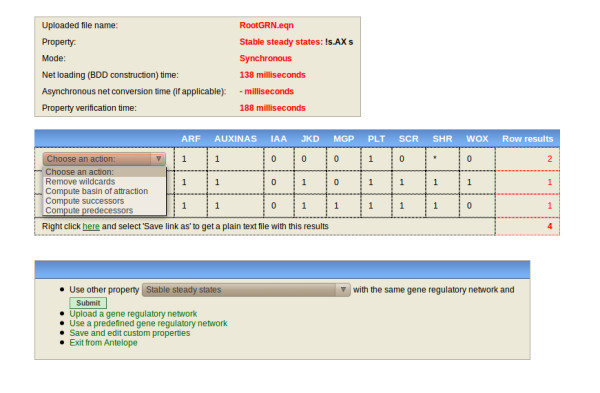
Results: We have developed Antelope ("Analysis of Networks through TEmporal-LOgic sPEcifications", http://turing.iimas.unam.mx:8080/AntelopeWEB/), a model checker for analyzing and constructing Boolean GRNs. Currently, software systems for Boolean GRNs use branching time almost exclusively for asynchrony. Antelope, by contrast, also uses branching time for incompletely specified behavior and environment interaction. We show the usefulness of modeling these two phenomena in the development of a Boolean GRN of the Arabidopsis thaliana root stem cell niche.There are two obstacles to a direct approach when applying model checking to Boolean GRN analysis. First, ordinary model checkers normally only verify whether or not a given set of model states has a given property. In comparison, a model checker for Boolean GRNs is preferable if it reports the set of states having a desired property. Second, for efficiency, the expressiveness of many model checkers is limited, resulting in the inability to express some interesting properties of Boolean GRNs.Antelope tries to overcome these two drawbacks: Apart from reporting the set of all states having a given property, our model checker can express, at the expense of efficiency, some properties that ordinary model checkers (e.g., NuSMV) cannot. This additional expressiveness is achieved by employing a logic extending the standard Computation-Tree Logic (CTL) with hybrid-logic operators.
Conclusions: We illustrate the advantages of Antelope when (a) modeling incomplete networks and environment interaction, (b) exhibiting the set of all states having a given property, and (c) representing Boolean GRN properties with hybrid CTL.
Figures
References
-
- Espinosa-Soto C, Padilla-Longoria P, Alvarez-Buylla E. A gene regulatory network model for cell-fate determination during Arabidopsis thaliana flower development that is robust and recovers experimental gene expression profiles. The Plant Cell. 2004;16:2923–2939. doi: 10.1105/tpc.104.021725. - DOI - PMC - PubMed
-
- Albert R. In: Complex Networks. Ben-Naim E, Frauenfelder H, Toroczkai Z, editor. Springer; 2004. Boolean modeling of genetic regulatory networks; pp. 459–481. [Lecture Notes in Physics Vol. 650]
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
