

A novel GRK2/HDAC6 interaction modulates cell spreading and motility
- PMID: 22193721
- PMCID: PMC3280560
- DOI: 10.1038/emboj.2011.466
A novel GRK2/HDAC6 interaction modulates cell spreading and motility
Abstract
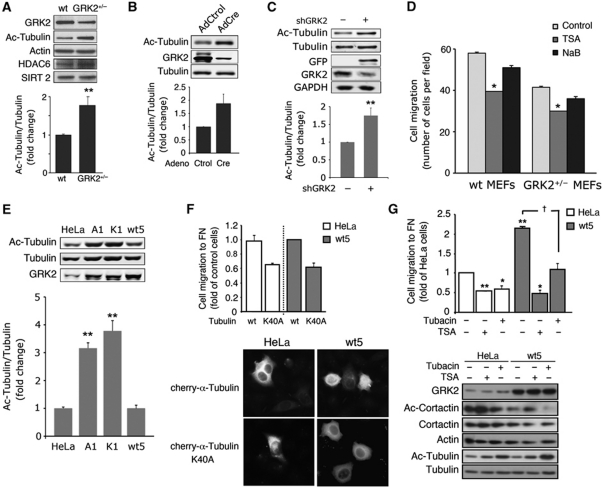
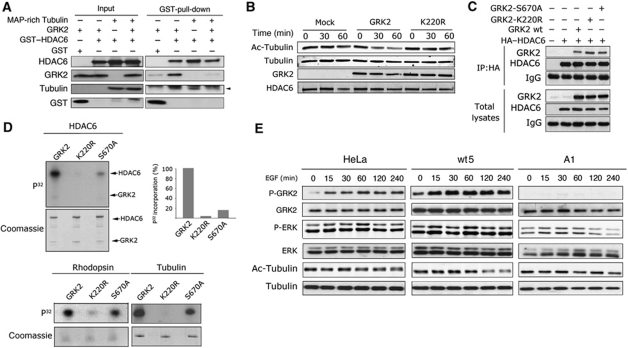
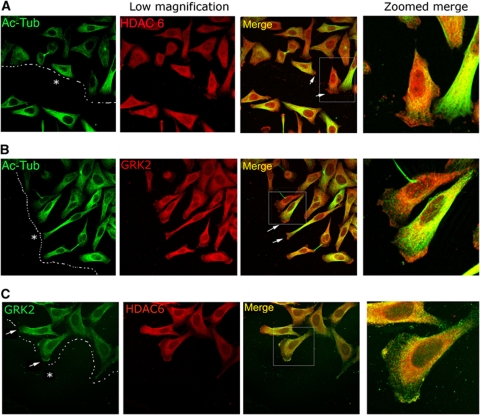
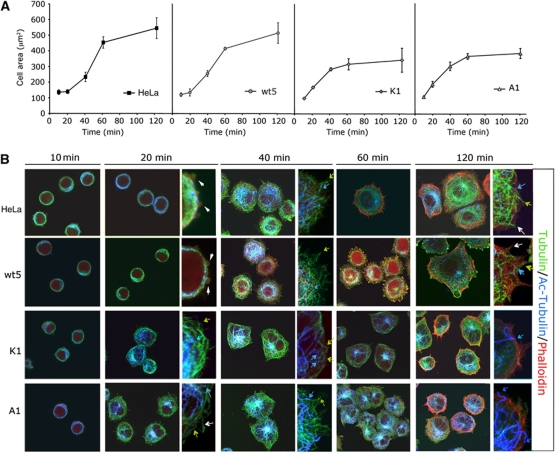
Cell motility and adhesion involves dynamic microtubule (MT) acetylation/deacetylation, a process regulated by enzymes as HDAC6, a major cytoplasmic α-tubulin deacetylase. We identify G protein-coupled receptor kinase 2 (GRK2) as a key novel stimulator of HDAC6. GRK2, which levels inversely correlate with the extent of α-tubulin acetylation in epithelial cells and fibroblasts, directly associates with and phosphorylates HDAC6 to stimulate α-tubulin deacetylase activity. Remarkably, phosphorylation of GRK2 itself at S670 specifically potentiates its ability to regulate HDAC6. GRK2 and HDAC6 colocalize in the lamellipodia of migrating cells, leading to local tubulin deacetylation and enhanced motility. Consistently, cells expressing GRK2-K220R or GRK2-S670A mutants, unable to phosphorylate HDAC6, exhibit highly acetylated cortical MTs and display impaired migration and protrusive activity. Finally, we find that a balanced, GRK2/HDAC6-mediated regulation of tubulin acetylation differentially modulates the early and late stages of cellular spreading. This novel GRK2/HDAC6 functional interaction may have important implications in pathological contexts.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




Similar articles
-
Roles of GRK2 in cell signaling beyond GPCR desensitization: GRK2-HDAC6 interaction modulates cell spreading and motility.Sci Signal. 2012 May 15;5(224):pt3. doi: 10.1126/scisignal.2003098. Sci Signal. 2012. PMID: 22589388
-
The interplay between G protein-coupled receptor kinase 2 (GRK2) and histone deacetylase 6 (HDAC6) at the crossroads of epithelial cell motility.Cell Adh Migr. 2012 Nov-Dec;6(6):495-501. doi: 10.4161/cam.21585. Epub 2012 Oct 17. Cell Adh Migr. 2012. PMID: 23076141 Free PMC article.
-
Extracellular signal-regulated kinase (ERK) phosphorylates histone deacetylase 6 (HDAC6) at serine 1035 to stimulate cell migration.J Biol Chem. 2013 Nov 15;288(46):33156-70. doi: 10.1074/jbc.M113.472506. Epub 2013 Oct 2. J Biol Chem. 2013. PMID: 24089523 Free PMC article.
-
Role of G protein-coupled receptor kinases in cell migration.Curr Opin Cell Biol. 2014 Apr;27:10-7. doi: 10.1016/j.ceb.2013.10.005. Epub 2013 Nov 16. Curr Opin Cell Biol. 2014. PMID: 24680425 Review.
-
HDAC6 α-tubulin deacetylase: a potential therapeutic target in neurodegenerative diseases.J Neurol Sci. 2011 May 15;304(1-2):1-8. doi: 10.1016/j.jns.2011.02.017. Epub 2011 Mar 5. J Neurol Sci. 2011. PMID: 21377170 Review.
Cited by
-
G protein-coupled receptor kinase 2 (GRK2) as a multifunctional signaling hub.Cell Mol Life Sci. 2019 Nov;76(22):4423-4446. doi: 10.1007/s00018-019-03274-3. Epub 2019 Aug 20. Cell Mol Life Sci. 2019. PMID: 31432234 Free PMC article. Review.
-
Human histone deacetylase 6 shows strong preference for tubulin dimers over assembled microtubules.Sci Rep. 2017 Sep 14;7(1):11547. doi: 10.1038/s41598-017-11739-3. Sci Rep. 2017. PMID: 28912522 Free PMC article.
-
Regulation of HDAC6 Catalytic Activity in Cancer: The Role of Post-Translational Modifications and Protein-Protein Interactions.Int J Mol Sci. 2025 Feb 1;26(3):1274. doi: 10.3390/ijms26031274. Int J Mol Sci. 2025. PMID: 39941046 Free PMC article. Review.
-
Histone deacetylase 6 in cancer.J Hematol Oncol. 2018 Sep 3;11(1):111. doi: 10.1186/s13045-018-0654-9. J Hematol Oncol. 2018. PMID: 30176876 Free PMC article. Review.
-
Histone deacetylase 6 is a FoxO transcription factor-dependent effector in skeletal muscle atrophy.J Biol Chem. 2015 Feb 13;290(7):4215-24. doi: 10.1074/jbc.M114.600916. Epub 2014 Dec 15. J Biol Chem. 2015. PMID: 25516595 Free PMC article.
References
-
- Azuma K, Urano T, Horie-Inoue K, Hayashi S, Sakai R, Ouchi Y, Inoue S (2009) Association of estrogen receptor alpha and histone deacetylase 6 causes rapid deacetylation of tubulin in breast cancer cells. Cancer Res 69: 2935–2940 - PubMed
-
- Boyault C, Sadoul K, Pabion M, Khochbin S (2007) HDAC6, at the crossroads between cytoskeleton and cell signaling by acetylation and ubiquitination. Oncogene 26: 5468–5476 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
