

Variation in mesoderm specification across Drosophilids is compensated by different rates of myoblast fusion during body wall musculature development
- PMID: 22194964
- PMCID: PMC3237579
- DOI: 10.1371/journal.pone.0028970
Variation in mesoderm specification across Drosophilids is compensated by different rates of myoblast fusion during body wall musculature development
Abstract
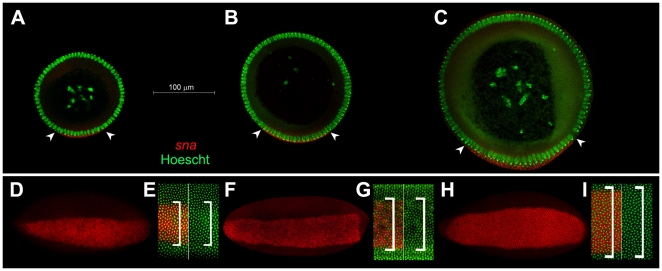
Background: It has been shown that species separated by relatively short evolutionary distances may have extreme variations in egg size and shape. Those variations are expected to modify the polarized morphogenetic gradients that pattern the dorso-ventral axis of embryos. Currently, little is known about the effects of scaling over the embryonic architecture of organisms. We began examining this problem by asking if changes in embryo size in closely related species of Drosophila modify all three dorso-ventral germ layers or only particular layers, and whether or not tissue patterning would be affected at later stages.
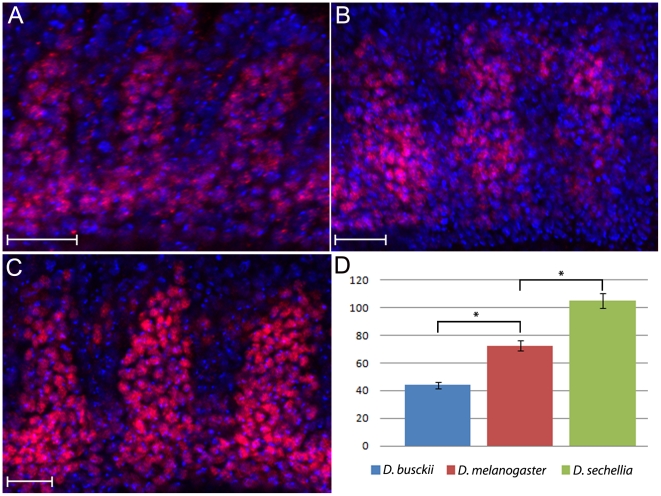
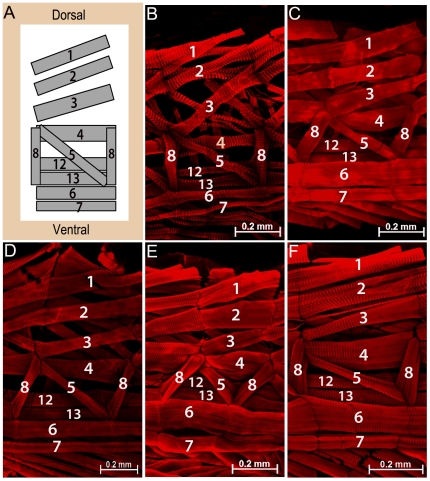
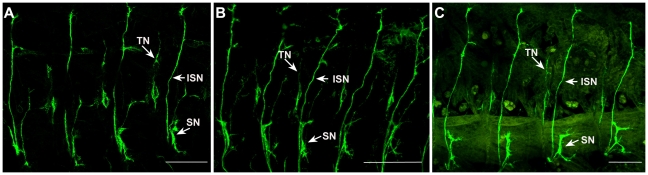
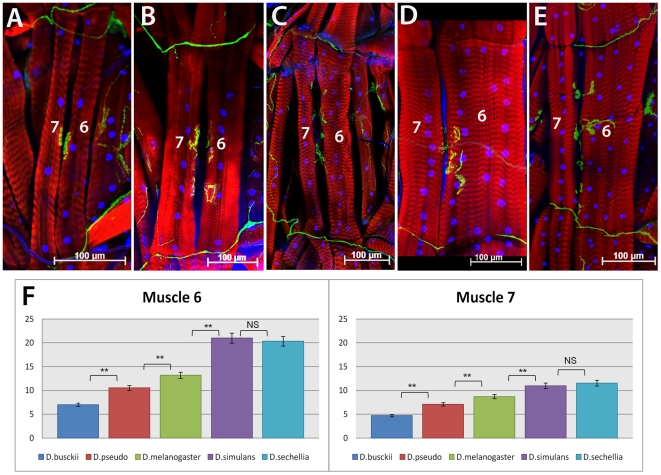
Principal findings: Here we report that changes in scale affect predominantly the mesodermal layer at early stages, while the neuroectoderm remains constant across the species studied. Next, we examined the fate of somatic myoblast precursor cells that derive from the mesoderm to test whether the assembly of the larval body wall musculature would be affected by the variation in mesoderm specification. Our results show that in all four species analyzed, the stereotyped organization of the body wall musculature is not disrupted and remains the same as in D. melanogaster. Instead, the excess or shortage of myoblast precursors is compensated by the formation of individual muscle fibers containing more or less fused myoblasts.
Conclusions: Our data suggest that changes in embryonic scaling often lead to expansions or retractions of the mesodermal domain across Drosophila species. At later stages, two compensatory cellular mechanisms assure the formation of a highly stereotyped larval somatic musculature: an invariable selection of 30 muscle founder cells per hemisegment, which seed the formation of a complete array of muscle fibers, and a variable rate in myoblast fusion that modifies the number of myoblasts that fuse to individual muscle fibers.
Conflict of interest statement
Figures

References
-
- Markow TA, Beall S, Matzkin LM. Egg size, embryonic development time and ovoviviparity in Drosophila species. J Evol Biol. 2009;22:430–434. - PubMed
-
- Irish VF, Gelbart WM. The decapentaplegic gene is required for dorsal-ventral patterning of the Drosophila embryo. Genes Dev. 1987;1:868–879. - PubMed
-
- Roth S, Stein D, Nusslein-Volhard C. A gradient of nuclear localization of the dorsal protein determines dorsoventral pattern in the Drosophila embryo. Cell. 1989;59:1189–1202. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
