

Nitric oxide signaling in the microcirculation
- PMID: 22196161
- PMCID: PMC3608675
- DOI: 10.1615/critrevbiomedeng.v39.i5.40
Nitric oxide signaling in the microcirculation
Abstract
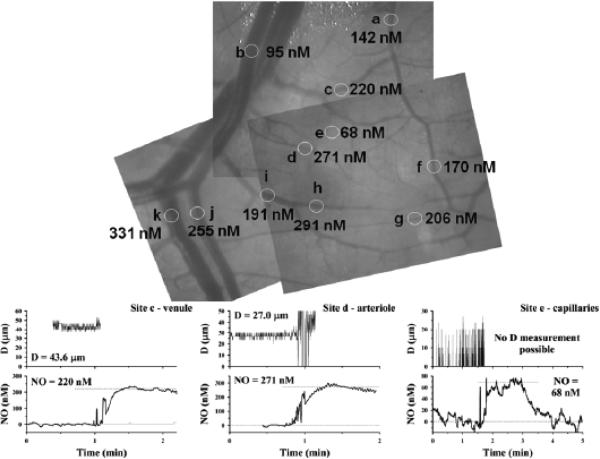
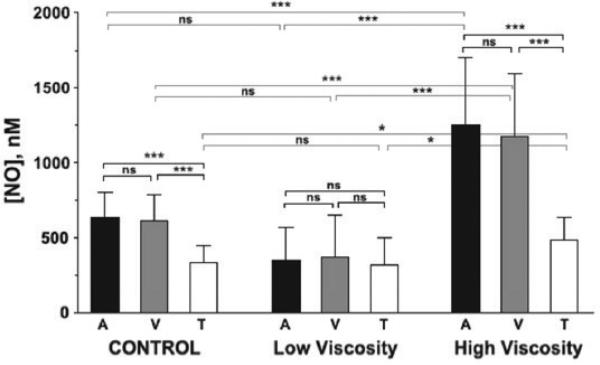
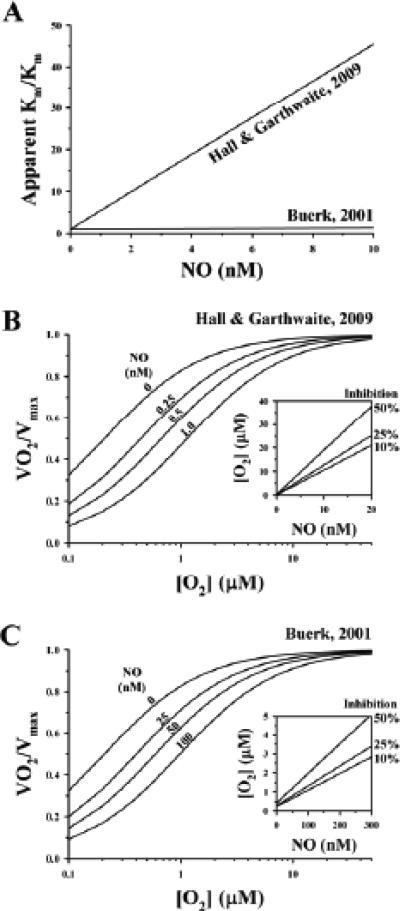
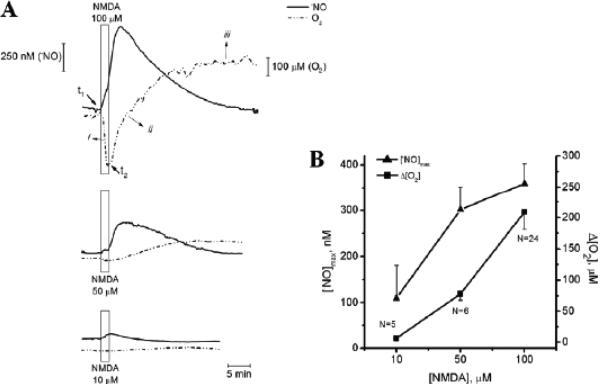
Several apparent paradoxes are evident when one compares mathematical predictions from models of nitric oxide (NO) diffusion and convection in vasculature structures with experimental measurements of NO (or related metabolites) in animal and human studies. Values for NO predicted from mathematical models are generally much lower than in vivo NO values reported in the literature for experiments, specifically with NO microelectrodes positioned at perivascular locations next to different sizes of blood vessels in the microcirculation and NO electrodes inserted into a wide range of tissues supplied by the microcirculation of each specific organ system under investigation. There continues to be uncertainty about the roles of NO scavenging by hemoglobin versus a storage function that may conserve NO, and other signaling targets for NO need to be considered. This review describes model predictions and relevant experimental data with respect to several signaling pathways in the microcirculation that involve NO.
Figures
References
-
- Murad F, Barber R. A hypothesis about cellular signaling with nitric oxide in the earliest life forms in evolution. Free Radic Biol Med. 2009;47(9):1325–7. - PubMed
-
- Palmer RM, Ferrige AG, Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;327(6122):524–6. - PubMed
-
- Wink DA, Hanbauer I, Grisham MB, Laval F, Nims RW, Laval J, et al. Chemical biology of nitric oxide: regulation and protective and toxic mechanisms. Curr Top Cell Regul. 1996;34:159–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources