

A poised chromatin platform for TGF-β access to master regulators
- PMID: 22196728
- PMCID: PMC3582033
- DOI: 10.1016/j.cell.2011.11.032
A poised chromatin platform for TGF-β access to master regulators
Abstract
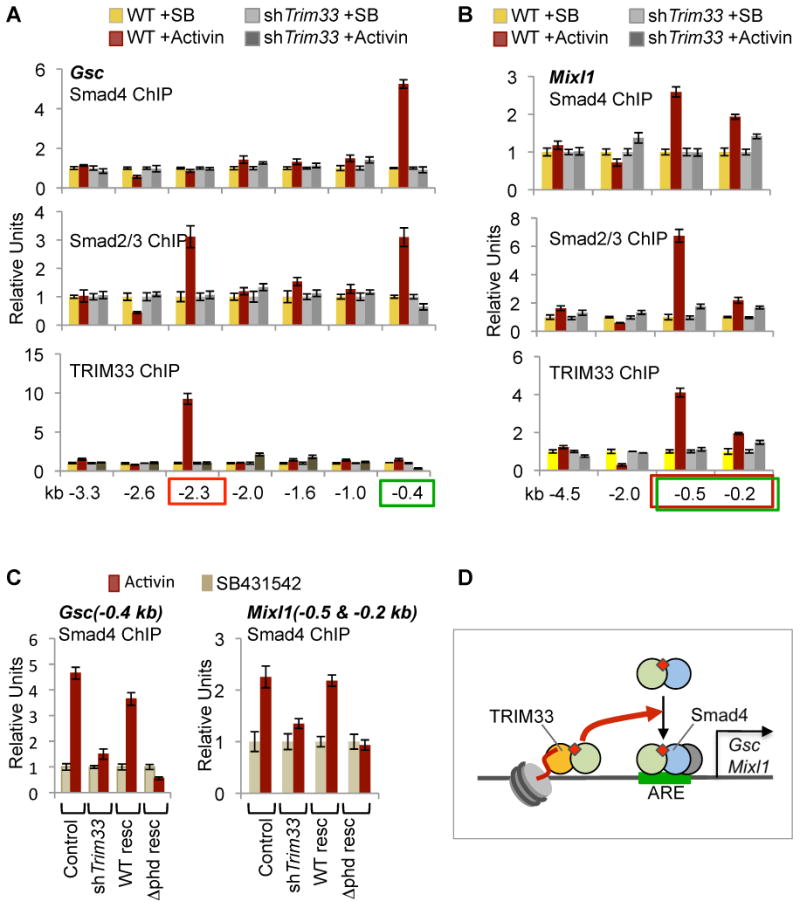
Specific chromatin marks keep master regulators of differentiation silent yet poised for activation by extracellular signals. We report that nodal TGF-β signals use the poised histone mark H3K9me3 to trigger differentiation of mammalian embryonic stem cells. Nodal receptors induce the formation of companion Smad4-Smad2/3 and TRIM33-Smad2/3 complexes. The PHD-Bromo cassette of TRIM33 facilitates binding of TRIM33-Smad2/3 to H3K9me3 and H3K18ac on the promoters of mesendoderm regulators Gsc and Mixl1. The crystal structure of this cassette, bound to histone H3 peptides, illustrates that PHD recognizes K9me3, and Bromo binds an adjacent K18ac. The interaction between TRIM33-Smad2/3 and H3K9me3 displaces the chromatin-compacting factor HP1γ, making nodal response elements accessible to Smad4-Smad2/3 for Pol II recruitment. In turn, Smad4 increases K18 acetylation to augment TRIM33-Smad2/3 binding. Thus, nodal effectors use the H3K9me3 mark as a platform to switch master regulators of stem cell differentiation from the poised to the active state.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures






References
-
- Agricola E, Randall RA, Gaarenstroom T, Dupont S, Hill CS. Recruitment of TIF1gamma to Chromatin via Its PHD Finger-Bromodomain Activates Its Ubiquitin Ligase and Transcriptional Repressor Activities. Mol Cell. 2011;43:85–96. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
