

Porcine Amelogenin : Alternative Splicing, Proteolytic Processing, Protein - Protein Interactions, and Possible Functions
- PMID: 22200995
- PMCID: PMC3245678
Porcine Amelogenin : Alternative Splicing, Proteolytic Processing, Protein - Protein Interactions, and Possible Functions
Abstract
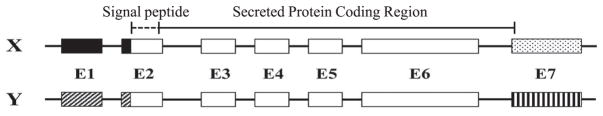
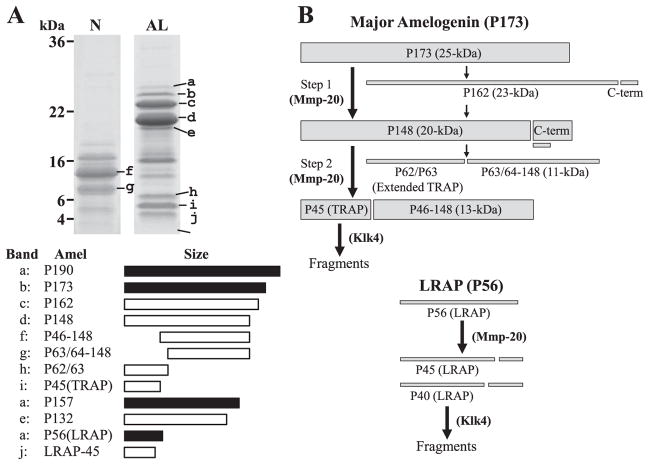
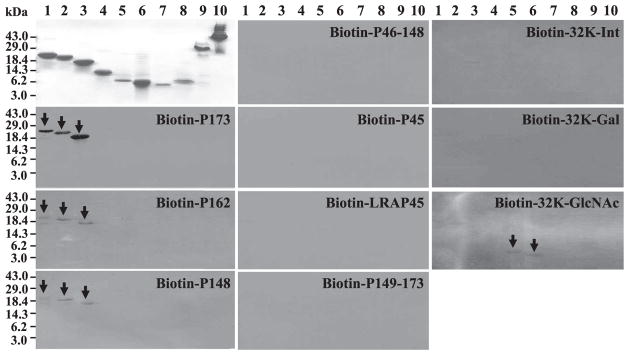
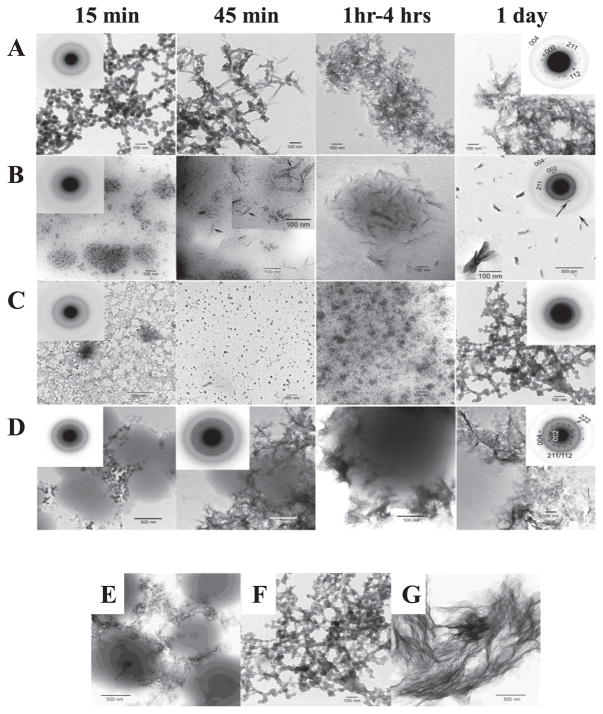
Amelogenin is the major secretory product of ameloblasts and is critical for proper tooth enamel formation. Amelogenin isoforms and their cleavage products comprise over 80% of total secretory stage enamel protein. We have isolated and characterized four secreted amelogenin isoforms from developing porcine enamel : P190 (27-kDa), P173 (25-kDa), P132 (18-kDa) and P56 (6.5-kDa ; leucine rich amelogenin polypeptide or LRAP). P190 and P132 are low abundance amelogenins that contain a novel exon 4-encoded segment of lack the exon 3-encoded segment, respectively. P173 is the most abundant (major) amelogenin isoform. Cleavage of P173 by matrix metalloproteinase 20 (Mmp20) occurs at specific sites that generates a set of N-terminal cleavage products : P162 (23-kDa), P148 (20-kDa), P62/P63 (11-kDa), and Trp(45) (6-kDa, tyrosine rich amelogenin polypeptide or TRAP). P148 is the most abundant protein in developing enamel and influences the conversion of amorphous calcium phosphate into hydroxyapatite in vitro. Mmp20 cleaves LRAP, the second abundant amelogenin isoform after Pro(45) and Pro(40). Processing by Mmp20 allows amelogenin cleavage products to serve separate functions. Over time, Mmp20 catalyzes additional cleavages that facilitate the progressive replacement of amelogenin by mineral, so enamel crystals thicken and widen with depth. Besides proteolytic processing, amelogenin protein-protein interactions are critical for function. Far-Western analyses demonstrate that the larger amelogenins (P173, P162, and P148) are only able to interact with larger amelogenins. No amelogenin-amelogenin interactions are observed for the smaller amelogenin cleavage products, TRAP or LRAP Amelogenin doesn't interact with the 32-kDa glycosylated enamelin cleavage product, unless it it partially deglycosylated.
Figures
Similar articles
-
MMP20 Proteolysis of Native Amelogenin Regulates Mineralization In Vitro.J Dent Res. 2016 Dec;95(13):1511-1517. doi: 10.1177/0022034516662814. Epub 2016 Aug 24. J Dent Res. 2016. PMID: 27558264 Free PMC article.
-
Mapping the Tooth Enamel Proteome and Amelogenin Phosphorylation Onto Mineralizing Porcine Tooth Crowns.Front Physiol. 2019 Jul 30;10:925. doi: 10.3389/fphys.2019.00925. eCollection 2019. Front Physiol. 2019. PMID: 31417410 Free PMC article.
-
Regulation of calcium phosphate formation by native amelogenins in vitro.Connect Tissue Res. 2014 Aug;55 Suppl 1(0):21-4. doi: 10.3109/03008207.2014.923853. Connect Tissue Res. 2014. PMID: 25158174 Free PMC article.
-
Expression, structure, and function of enamel proteinases.Connect Tissue Res. 2002;43(2-3):441-9. doi: 10.1080/03008200290001159. Connect Tissue Res. 2002. PMID: 12489196 Review.
-
Amelogenin proteins of developing dental enamel.Ciba Found Symp. 1997;205:118-30; discussion 130-4. doi: 10.1002/9780470515303.ch9. Ciba Found Symp. 1997. PMID: 9189621 Review.
Cited by
-
Tooth Enamel and its Dynamic Protein Matrix.Int J Mol Sci. 2020 Jun 23;21(12):4458. doi: 10.3390/ijms21124458. Int J Mol Sci. 2020. PMID: 32585904 Free PMC article. Review.
-
Exon4 amelogenin transcripts in enamel biomineralization.J Dent Res. 2015 Jun;94(6):836-42. doi: 10.1177/0022034515577412. Epub 2015 Mar 19. J Dent Res. 2015. PMID: 25792521 Free PMC article.
-
Dental enamel development: proteinases and their enamel matrix substrates.ISRN Dent. 2013 Sep 16;2013:684607. doi: 10.1155/2013/684607. ISRN Dent. 2013. PMID: 24159389 Free PMC article. Review.
-
A N-Terminus Domain Determines Amelogenin's Stability to Guide the Development of Mouse Enamel Matrix.J Bone Miner Res. 2021 Sep;36(9):1781-1795. doi: 10.1002/jbmr.4329. Epub 2021 May 25. J Bone Miner Res. 2021. PMID: 33957008 Free PMC article.
-
Phosphorylated and Non-phosphorylated Leucine Rich Amelogenin Peptide Differentially Affect Ameloblast Mineralization.Front Physiol. 2018 Feb 8;9:55. doi: 10.3389/fphys.2018.00055. eCollection 2018. Front Physiol. 2018. PMID: 29472869 Free PMC article.
References
-
- Uchida T, Tanabe T, Fukae M, Shimizu M, Yamada M, Miake K, Kobayashi S. Immunochemical and immunohistochemical studies, using antisera against porcine 25 kDa amelogenin, 89 kDa enamelin and the 13–17 kDa nonamelogenins, on immature enamel of the pig and rat. Histochemistry. 1991;96 :129–138. - PubMed
-
- Tanabe T, Fukae M, Uchida T, Shimizu M. The localization and characterization of proteinases for the initial cleavage of porcine amelogenin. Calcif Tissue Int. 1992;51 :213–217. - PubMed
-
- Nakahori Y, Takenaka O, Nakagome Y. A human X-Y homologous region encodes amelogenin. Genomics. 1991;9 :264–269. - PubMed
-
- Gibson CW, Golub E, Ding WD, Shimokawa H, Young M, Termine J, Rosenbloom J. Identification of the leucine-rich amelogenin peptide (LRAP) as the translation product of an alternatively spliced transcript. Biochem Biophys Res Commun. 1991;174 :1306–1312. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials