

Establishment of new mutations under divergence and genome hitchhiking
- PMID: 22201175
- PMCID: PMC3233718
- DOI: 10.1098/rstb.2011.0256
Establishment of new mutations under divergence and genome hitchhiking
Abstract
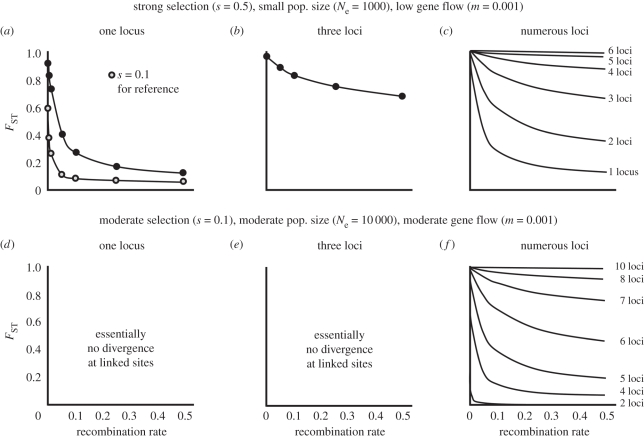
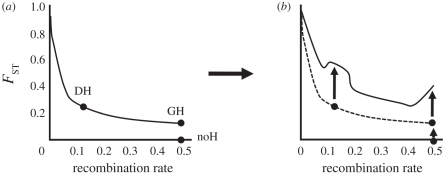
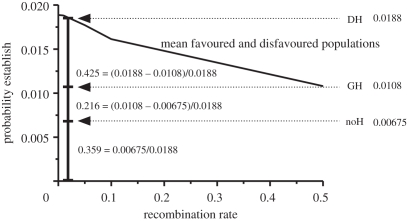
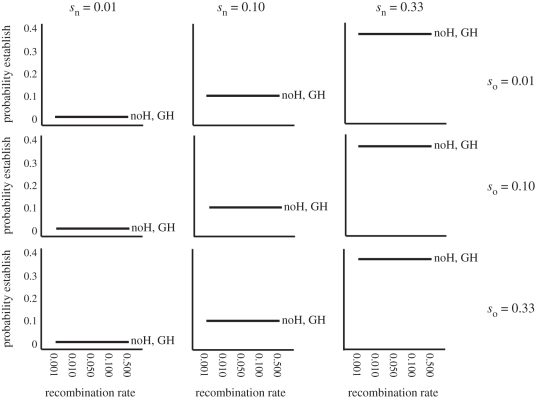
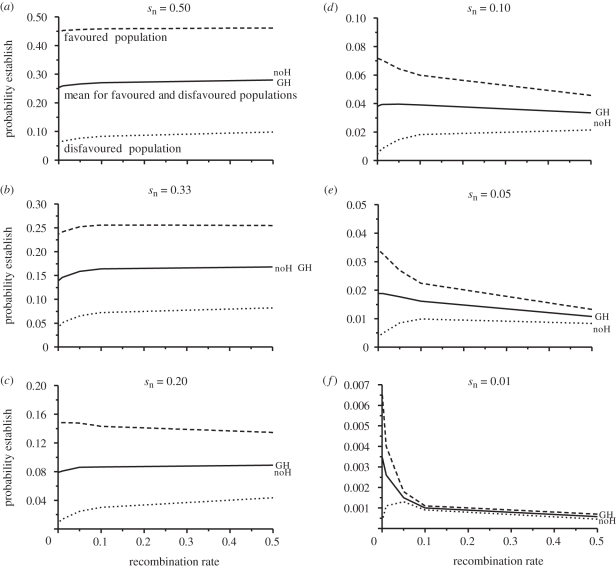
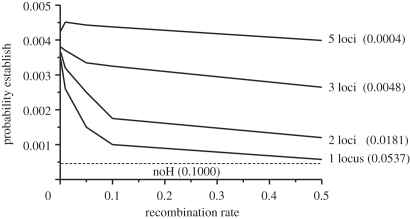
Theoretical models addressing genome-wide patterns of divergence during speciation are needed to help us understand the evolutionary processes generating empirical patterns. Here, we examine a critical issue concerning speciation-with-gene flow: to what degree does physical linkage (r < 0.5) of new mutations to already diverged genes aid the build-up of genomic islands of differentiation? We used simulation and analytical approaches to partition the probability of establishment for a new divergently selected mutation when the mutation (i) is the first to arise in an undifferentiated genome (the direct effect of selection), (ii) arises unlinked to any selected loci (r = 0.5), but within a genome that has some already diverged genes (the effect of genome-wide reductions in gene flow for facilitating divergence, which we term 'genome hitchhiking'), and (iii) arises in physical linkage to a diverged locus (divergence hitchhiking). We find that the strength of selection acting directly on a new mutation is generally the most important predictor for establishment, with divergence and genomic hitchhiking having smaller effects. We outline the specific conditions under which divergence and genome hitchhiking can aid mutation establishment. The results generate predictions about genome divergence at different points in the speciation process and avenues for further work.
Figures
Similar articles
-
The efficacy of divergence hitchhiking in generating genomic islands during ecological speciation.Evolution. 2010 Jun;64(6):1729-47. doi: 10.1111/j.1558-5646.2010.00943.x. Evolution. 2010. PMID: 20624183
-
Genetic hitchhiking and the dynamic buildup of genomic divergence during speciation with gene flow.Evolution. 2013 Sep;67(9):2577-91. doi: 10.1111/evo.12055. Epub 2013 Feb 12. Evolution. 2013. PMID: 24033168
-
A universal mechanism generating clusters of differentiated loci during divergence-with-migration.Evolution. 2016 Jul;70(7):1609-21. doi: 10.1111/evo.12957. Epub 2016 Jun 1. Evolution. 2016. PMID: 27196373 Free PMC article.
-
The genomics of speciation-with-gene-flow.Trends Genet. 2012 Jul;28(7):342-50. doi: 10.1016/j.tig.2012.03.009. Epub 2012 Apr 18. Trends Genet. 2012. PMID: 22520730 Review.
-
Divergent selection and heterogeneous genomic divergence.Mol Ecol. 2009 Feb;18(3):375-402. doi: 10.1111/j.1365-294X.2008.03946.x. Epub 2008 Dec 29. Mol Ecol. 2009. PMID: 19143936 Review.
Cited by
-
Incipient speciation between host-plant strains in the fall armyworm.BMC Ecol Evol. 2022 Apr 27;22(1):52. doi: 10.1186/s12862-022-02008-7. BMC Ecol Evol. 2022. PMID: 35477347 Free PMC article.
-
A major locus controls local adaptation and adaptive life history variation in a perennial plant.Genome Biol. 2018 Jun 4;19(1):72. doi: 10.1186/s13059-018-1444-y. Genome Biol. 2018. PMID: 29866176 Free PMC article.
-
Studying the genetic basis of speciation in high gene flow marine invertebrates.Curr Zool. 2016 Dec;62(6):643-653. doi: 10.1093/cz/zow093. Epub 2016 Aug 30. Curr Zool. 2016. PMID: 29491951 Free PMC article.
-
The effect of linkage on establishment and survival of locally beneficial mutations.Genetics. 2014 May;197(1):317-36. doi: 10.1534/genetics.114.163477. Epub 2014 Mar 7. Genetics. 2014. PMID: 24610861 Free PMC article.
-
Reproductive isolation via polygenic local adaptation in sub-divided populations: Effect of linkage disequilibria and drift.PLoS Genet. 2022 Sep 1;18(9):e1010297. doi: 10.1371/journal.pgen.1010297. eCollection 2022 Sep. PLoS Genet. 2022. PMID: 36048903 Free PMC article.
References
-
- Coyne J. A., Orr H. A. 2004. Speciation. Sunderland, MA: Sinauer Associates
-
- Mallet J. 1995. A species definition for the modern synthesis. Trends Ecol. Evol. 10, 294–29910.1016/0169-5347(95)90031-4 (doi:10.1016/0169-5347(95)90031-4) - DOI - DOI - PubMed
-
- Schluter D. 2001. Ecology and the origin of species. Trends Ecol. Evol. 16, 372–38010.1016/S0169-5347(01)02198-X (doi:10.1016/S0169-5347(01)02198-X) - DOI - DOI - PubMed
-
- Schluter D. 2009. Evidence for ecological speciation and its alternative. Science 323, 737–74110.1126/science.1160006 (doi:10.1126/science.1160006) - DOI - DOI - PubMed
-
- Rundle H. D., Nosil P. 2005. Ecological speciation. Ecol. Lett. 8, 336–35210.1111/j.1461-0248.2004.00715.x (doi:10.1111/j.1461-0248.2004.00715.x) - DOI - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
