

Mechanical compression drives cancer cells toward invasive phenotype
- PMID: 22203958
- PMCID: PMC3271885
- DOI: 10.1073/pnas.1118910109
Mechanical compression drives cancer cells toward invasive phenotype
Abstract
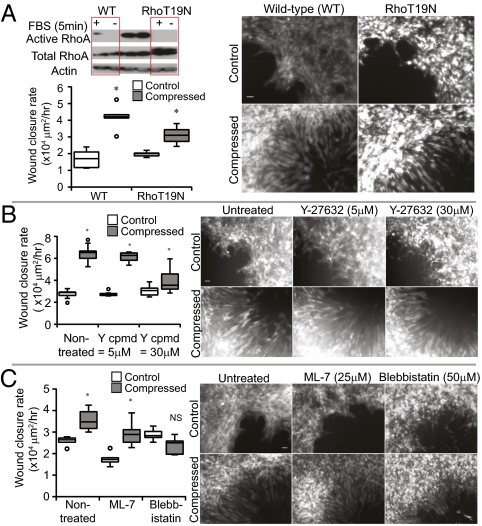
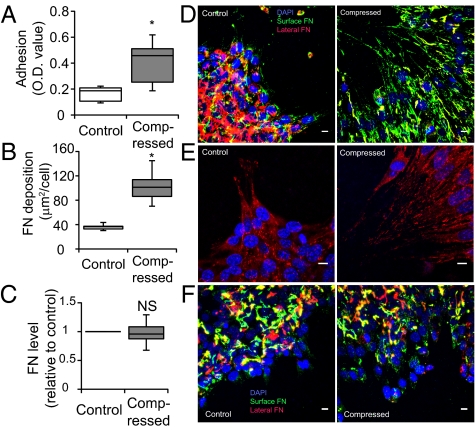
Uncontrolled growth in a confined space generates mechanical compressive stress within tumors, but little is known about how such stress affects tumor cell behavior. Here we show that compressive stress stimulates migration of mammary carcinoma cells. The enhanced migration is accomplished by a subset of "leader cells" that extend filopodia at the leading edge of the cell sheet. Formation of these leader cells is dependent on cell microorganization and is enhanced by compressive stress. Accompanied by fibronectin deposition and stronger cell-matrix adhesion, the transition to leader-cell phenotype results in stabilization of persistent actomyosin-independent cell extensions and coordinated migration. Our results suggest that compressive stress accumulated during tumor growth can enable coordinated migration of cancer cells by stimulating formation of leader cells and enhancing cell-substrate adhesion. This novel mechanism represents a potential target for the prevention of cancer cell migration and invasion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Fidler IJ, Poste G. The “seed and soil” hypothesis revisited. Lancet Oncol. 2008;9:808. - PubMed
-
- Wong SY, Hynes RO. Tumor-lymphatic interactions in an activated stromal microenvironment. J Cell Biochem. 2007;101:840–850. - PubMed
-
- Bierie B, Moses HL. Tumour microenvironment: TGFbeta: The molecular Jekyll and Hyde of cancer. Nat Rev Cancer. 2006;6:506–520. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
