

Geometry sensing by dendritic cells dictates spatial organization and PGE(2)-induced dissolution of podosomes
- PMID: 22204022
- PMCID: PMC3350765
- DOI: 10.1007/s00018-011-0908-y
Geometry sensing by dendritic cells dictates spatial organization and PGE(2)-induced dissolution of podosomes
Abstract
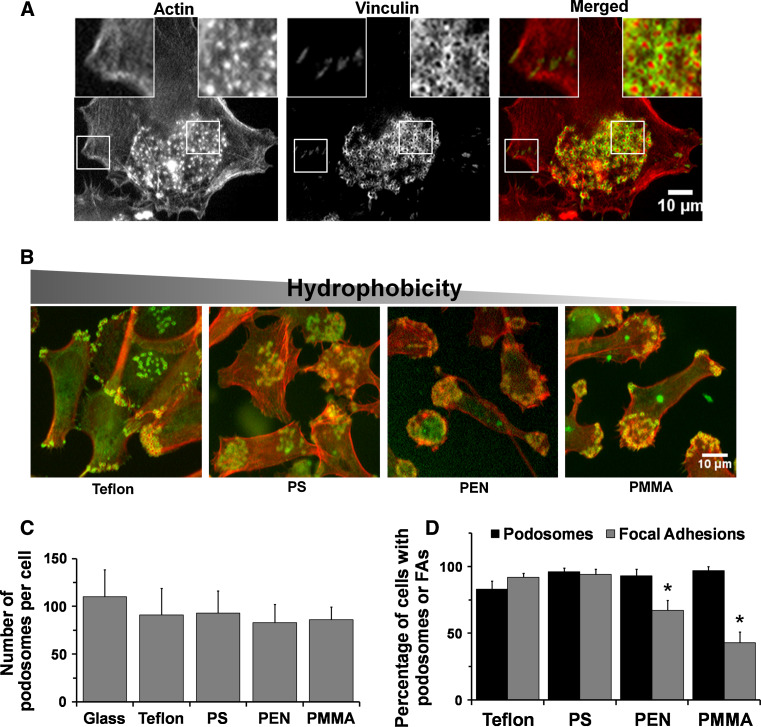
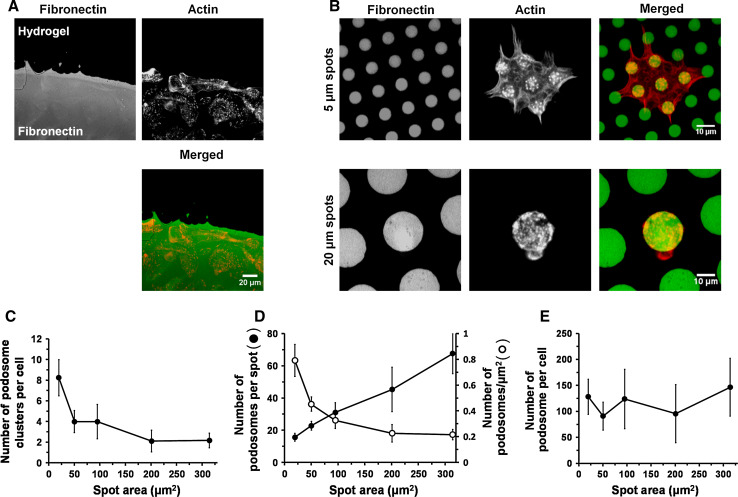
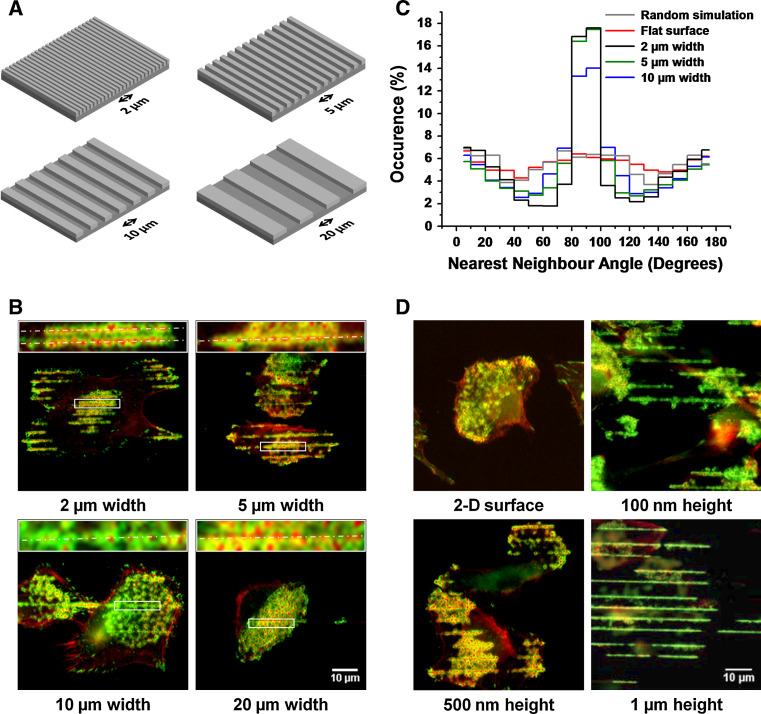
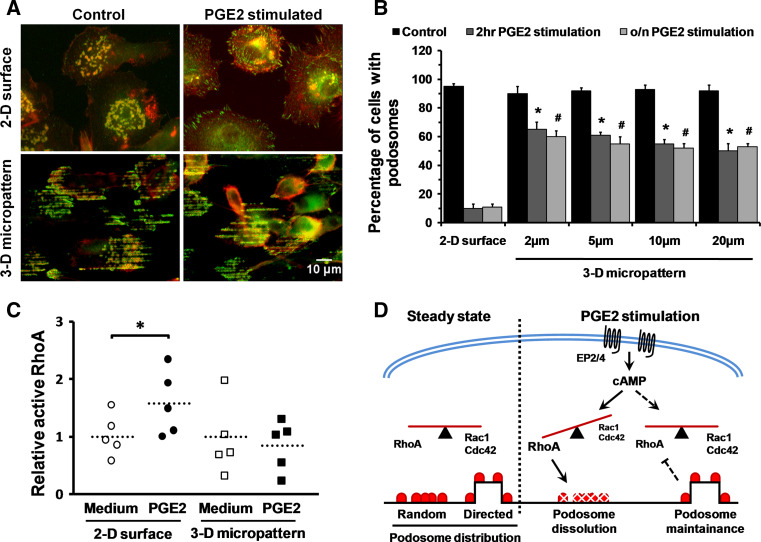
Assembly and disassembly of adhesion structures such as focal adhesions (FAs) and podosomes regulate cell adhesion and differentiation. On antigen-presenting dendritic cells (DCs), acquisition of a migratory and immunostimulatory phenotype depends on podosome dissolution by prostaglandin E(2) (PGE(2)). Whereas the effects of physico-chemical and topographical cues have been extensively studied on FAs, little is known about how podosomes respond to these signals. Here, we show that, unlike for FAs, podosome formation is not controlled by substrate physico-chemical properties. We demonstrate that cell adhesion is the only prerequisite for podosome formation and that substrate availability dictates podosome density. Interestingly, we show that DCs sense 3-dimensional (3-D) geometry by aligning podosomes along the edges of 3-D micropatterned surfaces. Finally, whereas on a 2-dimensional (2-D) surface PGE(2) causes a rapid increase in activated RhoA levels leading to fast podosome dissolution, 3-D geometric cues prevent PGE(2)-mediated RhoA activation resulting in impaired podosome dissolution even after prolonged stimulation. Our findings indicate that 2-D and 3-D geometric cues control the spatial organization of podosomes. More importantly, our studies demonstrate the importance of substrate dimensionality in regulating podosome dissolution and suggest that substrate dimensionality plays an important role in controlling DC activation, a key process in initiating immune responses.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
