

Review
doi: 10.1186/gb-2011-12-12-236.
Alu elements: know the SINEs
Affiliations
- PMID: 22204421
- PMCID: PMC3334610
- DOI: 10.1186/gb-2011-12-12-236
Item in Clipboard
Review
Alu elements: know the SINEs
Genome Biol.
.
Abstract
Alu elements are primate-specific repeats and comprise 11% of the human genome. They have wide-ranging influences on gene expression. Their contribution to genome evolution, gene regulation and disease is reviewed.
Figures

The structure of an Alu element. (a) The top portion shows a genomic Alu element between two direct repeats formed at the site of insertion (red arrowheads). The Alu ends with a long A-run, often referred to as the A-tail, and it also has a smaller A-rich region (indicated by AA) separating the two halves of a diverged dimer structure. Alu elements have the internal components of a RNA polymerase III promoter (boxes A and B), but they do not encode a terminator for RNA polymerase III. They utilize whatever stretch of T nucleotides is found at various distances downstream of the Alu element to terminate transcription. A typical Alu transcript is shown below the genomic Alu, showing that it encompasses the entire Alu, including the A-tail, and has a 3' region that is unique for each locus. (b) The Alu RNA is thought to fold into separate structures for each monomer unit. The RNA has been shown to bind the 7SL RNA SRP9 and 14 heterodimer, as well as polyA-binding protein (PABP). It is thought that at least one other protein binds the duplex portion of the RNA structure. (c) In the target-primed reverse transcription mechanism, the Alu RNA (blue) brings the ORF2p to the genome where its endonuclease activity cleaves at a T-rich consensus sequence. The T-rich region primes reverse transcription by ORF2p on the 3' A-tail region of the Alu element. This creates a cDNA copy of the body of the Alu element. A nick occurs by an unknown mechanism on the second strand and second-strand synthesis is primed. The new Alu element is then flanked by short direct repeats that are duplicates of the DNA sequence between the first and second nicks.

Why so few Alu elements are active. Out of the more than 1 million Alu elements in the human genome, very few are capable of making copies, although many make transcripts. Upon insertion in a new locus, the factors that make a very active Alu element are the flanking sequences influencing the promoter, creating a short unique region. Active elements match the consensus Alu element fairly closely and they have a long and fairly perfect A-tail. Active elements degrade rapidly on an evolutionary time scale by A-tail shortening, heterogeneous base interruptions accumulating in the A-tail, and eventually by the accumulation of random mutations in the Alu element. At least some of these changes alter Alu activity through disruption of the various proteins binding to the RNA in the ribonucleoprotein (Figure 1b).
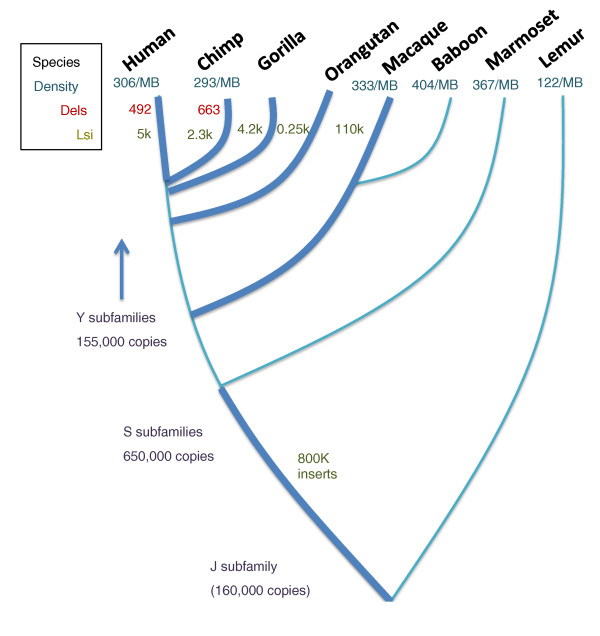
Evolutionary impact of Alu elements in primates. An approximate evolutionary tree is shown for various primate species. The approximate density of Alu elements in the genomes of those species is shown as the number of Alu elements per megabase (MB). For specific evolutionary time periods, marked by thicker lines, the number of lineage-specific Alu insertions (Lsi) is marked. Data of Alu/Alu recombination causing deletions (Dels) between the human and chimp genomes are also shown. Note that the rate of Alu insertion, as well as recombination, seems to vary with different lineages and different evolutionary time periods.
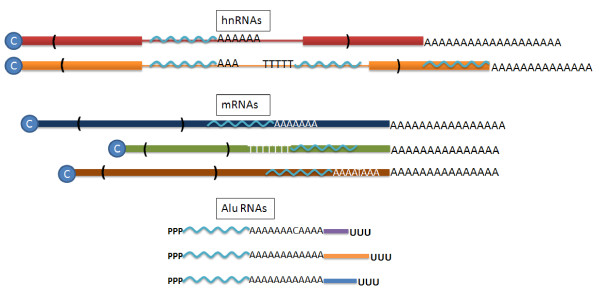
RNAs containing Alu sequences. Because of the high density of Alu elements found in introns, there are many Alu elements located (in both sense and antisense orientations) within the introns of the primary nuclear transcripts (hnRNAs) found in the nucleus. Even after splicing, a high proportion of mature cytoplasmic mRNAs also contain Alu elements in their 3' non-coding regions. Both of the above types of transcript involve capped (blue circle with C), polyadenylated RNAs where one or more Alu elements is included in either orientation within the transcript. The transcripts made from the Alu RNA polymerase III promoter are not capped, include a genomically encoded A-rich region, and terminate at a typical RNA pol III terminator. The transcripts are similar, but not identical, in the Alu region (wavy light blue line), with an A-tail encoded from the genome that is variable in length and often includes non-A bases. The 3' ends of each Alu RNA are unique, and arise from the flanking sequences of the different genomic Alu elements (Figure 1).

Alu elements and post-transcriptional processing of transcripts. (a) The majority of primary transcripts from genes contain Alu elements, both sense and antisense, within their introns. These Alu elements gradually accumulate mutations that can activate cryptic splice sites, or polyadenylation sites, within the Alu. This can lead to alternative splicing of RNAs that can either include a portion of an Alu in the coding region or result in premature termination of translation. Similarly, Alu elements may cause premature termination and polyadenylation resulting in truncated genes. (b) Alu elements in introns located in opposite orientations can fold into secondary structures that are then a major substrate for ADAR (adenosine deaminase that acts on RNA) activity. The edited RNAs may then have cryptic splice sites activated or may also result in retention of the RNA in the nucleus.
Similar articles
-
True Homoplasy of Retrotransposon Insertions in Primates.Syst Biol. 2019 May 1;68(3):482-493. doi: 10.1093/sysbio/syy076. Syst Biol. 2019. PMID: 30445649
-
Alu retrotransposition-mediated deletion.J Mol Biol. 2005 May 13;348(4):791-800. doi: 10.1016/j.jmb.2005.02.043. J Mol Biol. 2005. PMID: 15843013
-
Non-traditional Alu evolution and primate genomic diversity.J Mol Biol. 2002 Mar 8;316(5):1033-40. doi: 10.1006/jmbi.2001.5380. J Mol Biol. 2002. PMID: 11884141
-
Alu repeats and human genomic diversity.Nat Rev Genet. 2002 May;3(5):370-9. doi: 10.1038/nrg798. Nat Rev Genet. 2002. PMID: 11988762 Review.
-
[Alu elements in the human genome].Tidsskr Nor Laegeforen. 2004 Sep 23;124(18):2345-9. Tidsskr Nor Laegeforen. 2004. PMID: 15467796 Review. Norwegian.
Cited by
-
A study of transposable element-associated structural variations (TASVs) using a de novo-assembled Korean genome.Exp Mol Med. 2021 Apr;53(4):615-630. doi: 10.1038/s12276-021-00586-y. Epub 2021 Apr 8. Exp Mol Med. 2021. PMID: 33833373 Free PMC article.
-
High-Throughput Analysis of Global DNA Methylation Using Methyl-Sensitive Digestion.PLoS One. 2016 Oct 17;11(10):e0163184. doi: 10.1371/journal.pone.0163184. eCollection 2016. PLoS One. 2016. PMID: 27749902 Free PMC article.
-
RNAs as Regulators of Cellular Matchmaking.Front Mol Biosci. 2021 Apr 9;8:634146. doi: 10.3389/fmolb.2021.634146. eCollection 2021. Front Mol Biosci. 2021. PMID: 33898516 Free PMC article. Review.
-
Recent insights into crosstalk between genetic parasites and their host genome.Brief Funct Genomics. 2024 Jan 18;23(1):15-23. doi: 10.1093/bfgp/elac032. Brief Funct Genomics. 2024. PMID: 36307128 Free PMC article. Review.
-
A reference genome for the critically endangered woylie, Bettongia penicillata ogilbyi.GigaByte. 2021 Dec 10;2021:gigabyte35. doi: 10.46471/gigabyte.35. eCollection 2021. GigaByte. 2021. PMID: 36824341 Free PMC article.
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, Funke R, Gage D, Harris K, Heaford A, Howland J, Kann L, Lehoczky J, LeVine R, McEwan P, McKernan K, Meldrim J, Mesirov JP, Miranda C, Morris W, Naylor J, Raymond C, Rosetti M, Santos R, Sheridan A, Sougnez C. et al.Initial sequencing and analysis of the human genome. International Human Genome Sequencing Consortium. Nature. 2001;409:860–921. doi: 10.1038/35057062. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
