

Self-assembly of viral capsid protein and RNA molecules of different sizes: requirement for a specific high protein/RNA mass ratio
- PMID: 22205731
- PMCID: PMC3302347
- DOI: 10.1128/JVI.06566-11
Self-assembly of viral capsid protein and RNA molecules of different sizes: requirement for a specific high protein/RNA mass ratio
Erratum in
-
Correction for Cadena-Nava et al., "Self-Assembly of Viral Capsid Protein and RNA Molecules of Different Sizes: Requirement for a Specific High Protein/RNA Mass Ratio".J Virol. 2019 Feb 5;93(4):e01939-18. doi: 10.1128/JVI.01939-18. Print 2019 Feb 15. J Virol. 2019. PMID: 30723138 Free PMC article. No abstract available.
Abstract
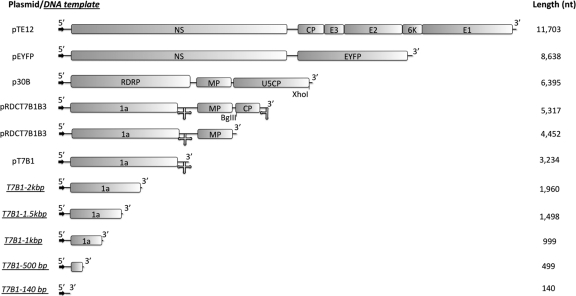
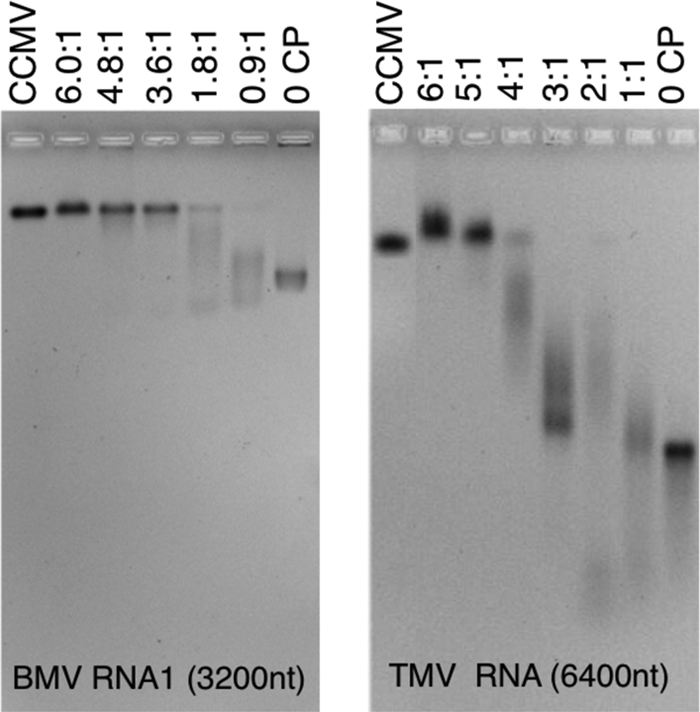
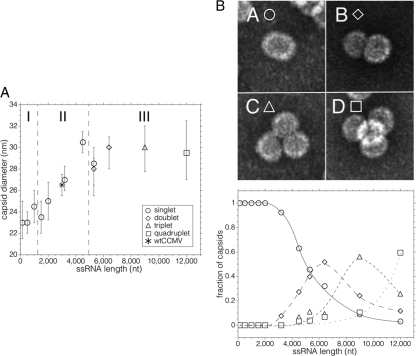
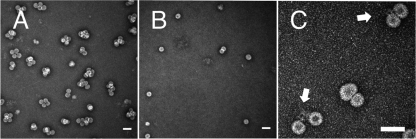
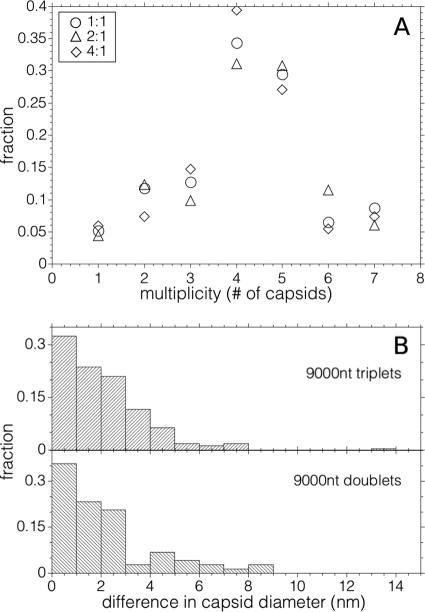
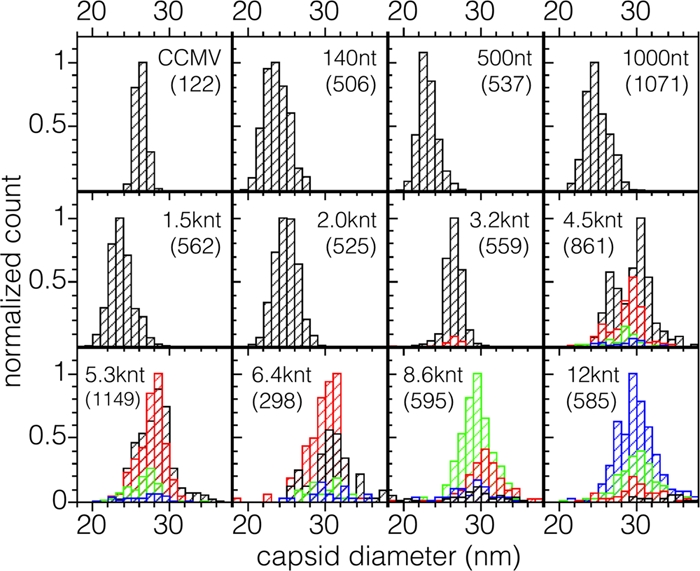
Virus-like particles can be formed by self-assembly of capsid protein (CP) with RNA molecules of increasing length. If the protein "insisted" on a single radius of curvature, the capsids would be identical in size, independent of RNA length. However, there would be a limit to length of the RNA, and one would not expect RNA much shorter than native viral RNA to be packaged unless multiple copies were packaged. On the other hand, if the protein did not favor predetermined capsid size, one would expect the capsid diameter to increase with increase in RNA length. Here we examine the self-assembly of CP from cowpea chlorotic mottle virus with RNA molecules ranging in length from 140 to 12,000 nucleotides (nt). Each of these RNAs is completely packaged if and only if the protein/RNA mass ratio is sufficiently high; this critical value is the same for all of the RNAs and corresponds to equal RNA and N-terminal-protein charges in the assembly mix. For RNAs much shorter in length than the 3,000 nt of the viral RNA, two or more molecules are assembled into 24- and 26-nm-diameter capsids, whereas for much longer RNAs (>4,500 nt), a single RNA molecule is shared/packaged by two or more capsids with diameters as large as 30 nm. For intermediate lengths, a single RNA is assembled into 26-nm-diameter capsids, the size associated with T=3 wild-type virus. The significance of these assembly results is discussed in relation to likely factors that maintain T=3 symmetry in vivo.
Figures

References
-
- Adolph KW, Butler PJG. 1977. Studies on the assembly of a spherical plant virus. III. Reassembly of infectious virions under mild conditions. J. Mol. Biol. 109:345–357 - PubMed
-
- Annamalai P, Rao ALN. 2005. Dispensability of 3′ tRNA-like sequence for packaging cowpea chlorotic mottle virus genomic RNAs. Virology 332:650–658 - PubMed
-
- Annamalai P, Rofail F, DeMason DA, Rao ALN. 2008. Replication-coupled packaging mechanism in positive-strand RNA viruses: synchronized coexpression of functional multigenome RNA components of an animal and a plant virus in Nicotiana benthamiana cells by agroinfiltration. J. Virol. 82:1484–1490 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
