

Mechanisms of MEOX1 and MEOX2 regulation of the cyclin dependent kinase inhibitors p21 and p16 in vascular endothelial cells
- PMID: 22206000
- PMCID: PMC3243699
- DOI: 10.1371/journal.pone.0029099
Mechanisms of MEOX1 and MEOX2 regulation of the cyclin dependent kinase inhibitors p21 and p16 in vascular endothelial cells
Abstract
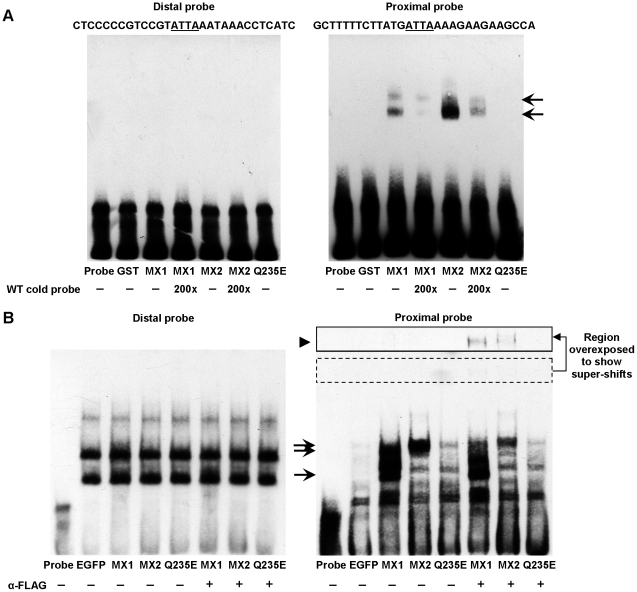
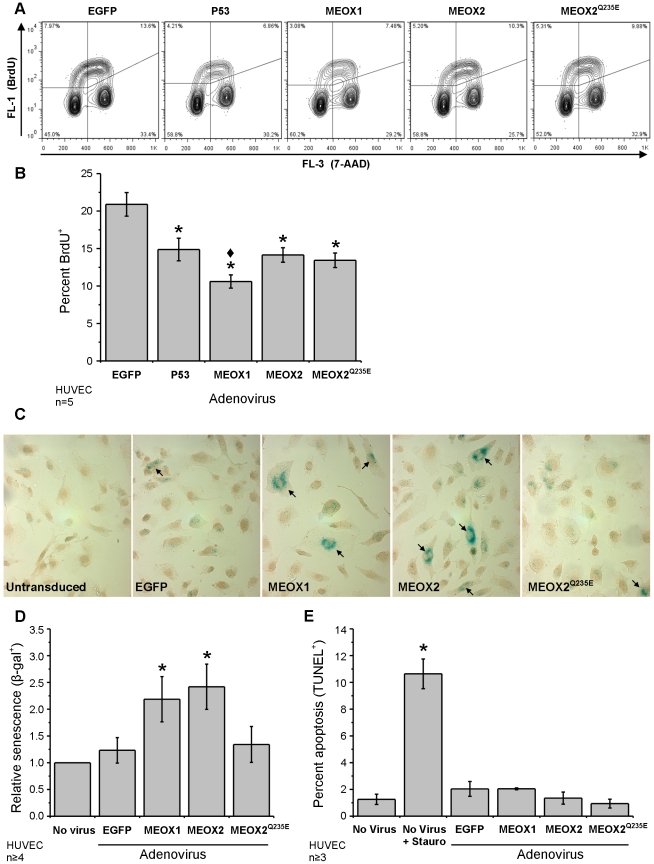
Senescence, the state of permanent cell cycle arrest, has been associated with endothelial cell dysfunction and atherosclerosis. The cyclin dependent kinase inhibitors p21(CIP1/WAF1) and p16(INK4a) govern the G(1)/S cell cycle checkpoint and are essential for determining whether a cell enters into an arrested state. The homeodomain transcription factor MEOX2 is an important regulator of vascular cell proliferation and is a direct transcriptional activator of both p21(CIP1/WAF1) and p16(INK4a). MEOX1 and MEOX2 have been shown to be partially functionally redundant during development, suggesting that they regulate similar target genes in vivo. We compared the ability of MEOX1 and MEOX2 to activate p21(CIP1/WAF1) and p16(INK4a) expression and induce endothelial cell cycle arrest. Our results demonstrate for the first time that MEOX1 regulates the MEOX2 target genes p21(CIP1/WAF1) and p16(INK4a). In addition, increased expression of either of the MEOX homeodomain transcription factors leads to cell cycle arrest and endothelial cell senescence. Furthermore, we show that the mechanism of transcriptional activation of these cyclin dependent kinase inhibitor genes by MEOX1 and MEOX2 is distinct. MEOX1 and MEOX2 activate p16(INK4a) in a DNA binding dependent manner, whereas they induce p21(CIP1/WAF1) in a DNA binding independent manner.
Conflict of interest statement
Figures





References
-
- Li WW, Talcott KE, Zhai AW, Kruger EA, Li VW. The role of therapeutic angiogenesis in tissue repair and regeneration. Adv Skin Wound Care. 2005;18:491–500; quiz 501-492. - PubMed
-
- Smith SK. Regulation of angiogenesis in the endometrium. Trends Endocrinol Metab. 2001;12:147–151. - PubMed
-
- Bloor CM. Angiogenesis during exercise and training. Angiogenesis. 2005;8:263–271. - PubMed
-
- Rivard A, Fabre JE, Silver M, Chen D, Murohara T, et al. Age-dependent impairment of angiogenesis. Circulation. 1999;99:111–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
