

Mitochondrial respiratory capacity is a critical regulator of CD8+ T cell memory development
- PMID: 22206904
- PMCID: PMC3269311
- DOI: 10.1016/j.immuni.2011.12.007
Mitochondrial respiratory capacity is a critical regulator of CD8+ T cell memory development
Abstract
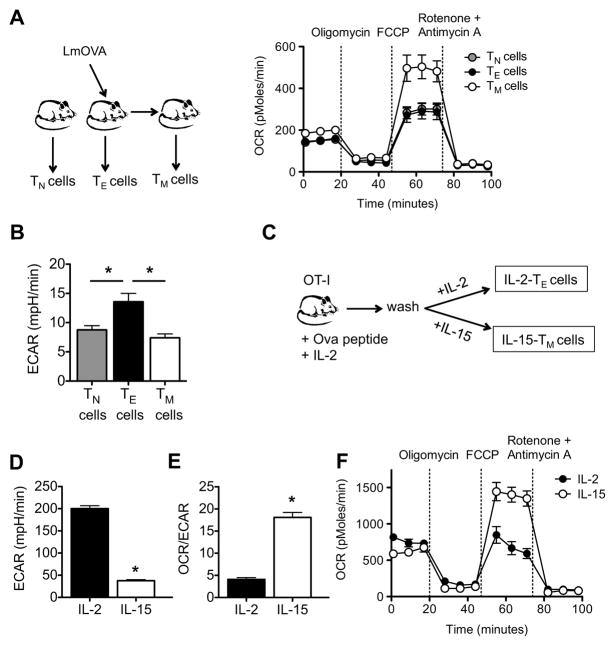
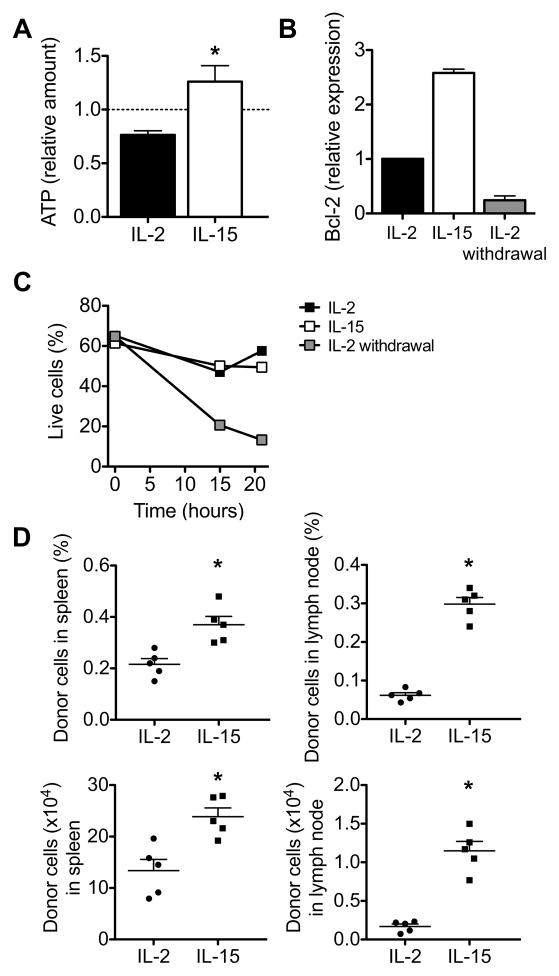
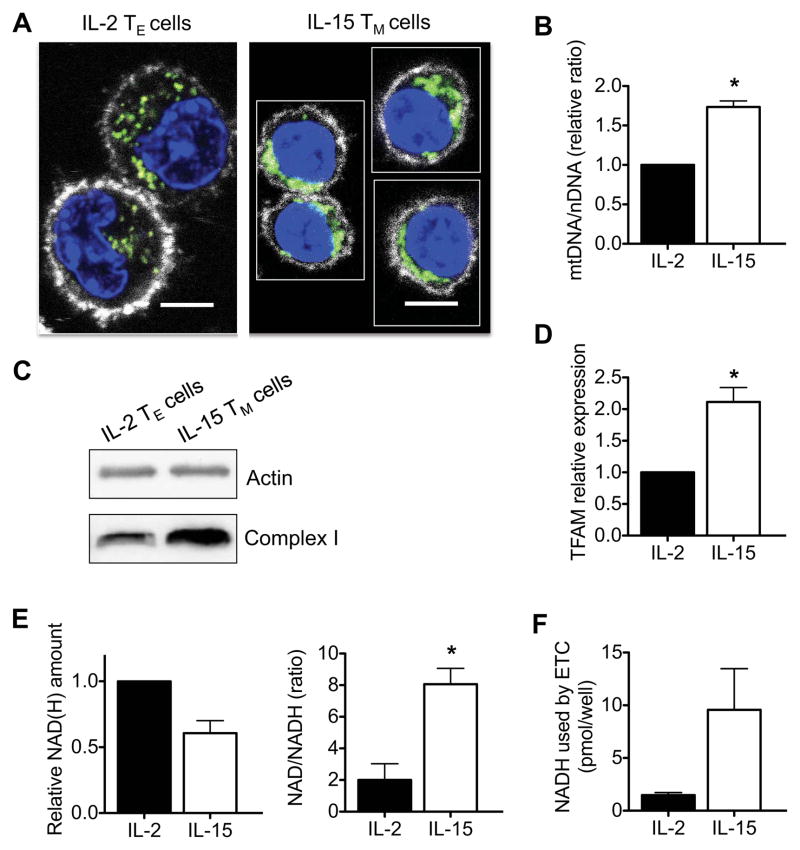
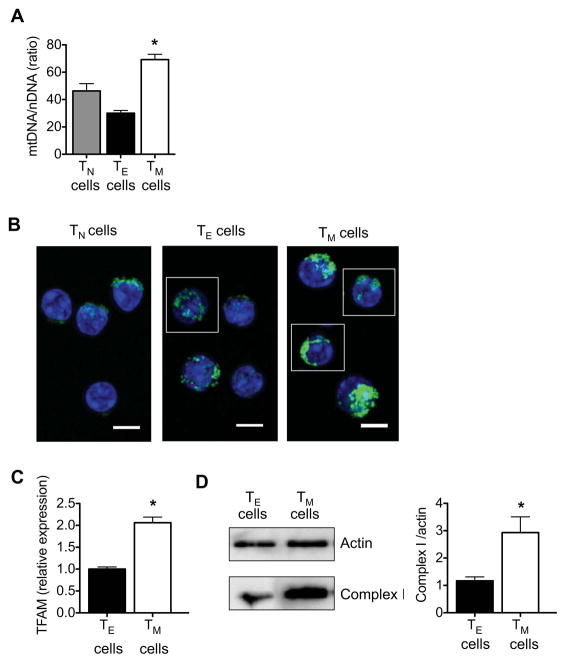
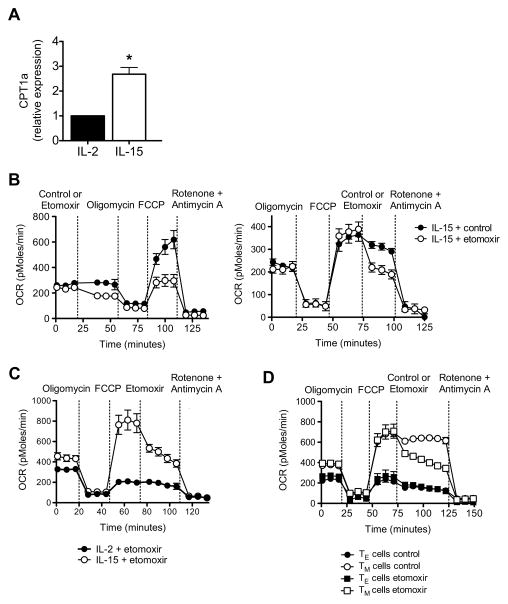
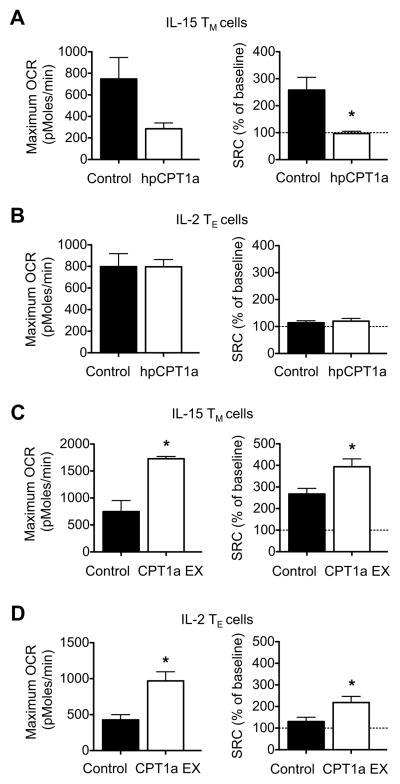
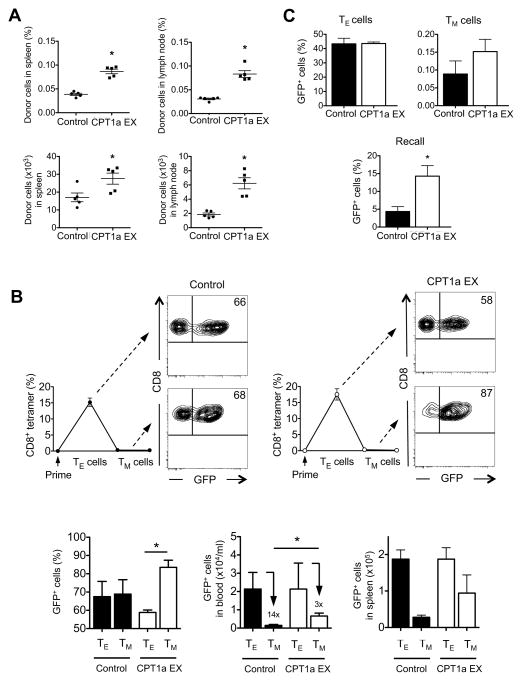
CD8(+) T cells undergo major metabolic changes upon activation, but how metabolism influences the establishment of long-lived memory T cells after infection remains a key question. We have shown here that CD8(+) memory T cells, but not CD8(+) T effector (Teff) cells, possessed substantial mitochondrial spare respiratory capacity (SRC). SRC is the extra capacity available in cells to produce energy in response to increased stress or work and as such is associated with cellular survival. We found that interleukin-15 (IL-15), a cytokine critical for CD8(+) memory T cells, regulated SRC and oxidative metabolism by promoting mitochondrial biogenesis and expression of carnitine palmitoyl transferase (CPT1a), a metabolic enzyme that controls the rate-limiting step to mitochondrial fatty acid oxidation (FAO). These results show how cytokines control the bioenergetic stability of memory T cells after infection by regulating mitochondrial metabolism.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Immunometabolism: IL-15 provides breathing space for memory.Nat Rev Immunol. 2012 Jan 25;12(2):76. doi: 10.1038/nri3162. Nat Rev Immunol. 2012. PMID: 22273770 No abstract available.
-
Fueling memories.Immunity. 2012 Jan 27;36(1):3-5. doi: 10.1016/j.immuni.2012.01.003. Immunity. 2012. PMID: 22284413 Free PMC article.
References
-
- Brand KA, Hermfisse U. Aerobic glycolysis by proliferating cells: a protective strategy against reactive oxygen species. FASEB J. 1997;11:388–395. - PubMed
-
- Carrio R, Bathe OF, Malek TR. Initial antigen encounter programs CD8+ T cells competent to develop into memory cells that are activated in an antigen-free, IL-7- and IL-15-rich environment. J Immunol. 2004;172:7315–7323. - PubMed
-
- Chang JT, Palanivel VR, Kinjyo I, Schambach F, Intlekofer AM, Banerjee A, Longworth SA, Vinup KE, Mrass P, Oliaro J, et al. Asymmetric T lymphocyte division in the initiation of adaptive immune responses. Science. 2007;315:1687–1691. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
