

Laws of conservation as related to brain growth, aging, and evolution: symmetry of the minicolumn
- PMID: 22207838
- PMCID: PMC3245969
- DOI: 10.3389/fnana.2011.00066
Laws of conservation as related to brain growth, aging, and evolution: symmetry of the minicolumn
Abstract
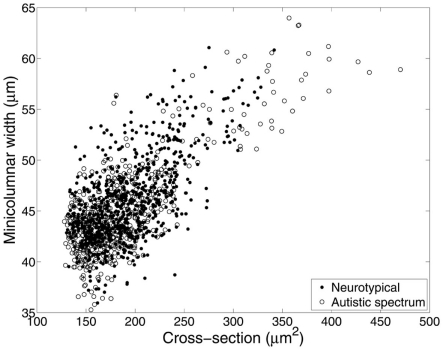
Development, aging, and evolution offer different time scales regarding possible anatomical transformations of the brain. This article expands on the perspective that the cerebral cortex exhibits a modular architecture with invariant properties in regards to these time scales. These properties arise from morphometric relations of the ontogenetic minicolumn as expressed in Noether's first theorem, i.e., that for each continuous symmetry there is a conserved quantity. Whenever minicolumnar symmetry is disturbed by either developmental or aging processes the principle of least action limits the scope of morphometric alterations. Alternatively, local and global divergences from these laws apply to acquired processes when the system is no longer isolated from its environment. The underlying precepts to these physical laws can be expressed in terms of mathematical equations that are conservative of quantity. Invariant properties of the brain include the rotational symmetry of minicolumns, a scaling proportion or "even expansion" between pyramidal cells and core minicolumnar size, and the translation of neuronal elements from the main axis of the minicolumn. It is our belief that a significant portion of the architectural complexity of the cerebral cortex, its response to injury, and its evolutionary transformation, can all be captured by a small set of basic physical laws dictated by the symmetry of minicolumns. The putative preservations of parameters related to the symmetry of the minicolumn suggest that the development and final organization of the cortex follows a deterministic process.
Keywords: cerebral cortex; minicolumns; neocortex; symmetry.
Figures
Similar articles
-
Noether's Theorem as a Metaphor for Chargaff's 2nd Parity Rule in Genomics.J Mol Evol. 2022 Aug;90(3-4):231-238. doi: 10.1007/s00239-022-10062-4. Epub 2022 Jun 15. J Mol Evol. 2022. PMID: 35704064
-
The Concept of Symmetry and the Theory of Perception.Front Comput Neurosci. 2021 Aug 23;15:681162. doi: 10.3389/fncom.2021.681162. eCollection 2021. Front Comput Neurosci. 2021. PMID: 34497499 Free PMC article.
-
Minicolumn size and human cortex.Prog Brain Res. 2012;195:219-35. doi: 10.1016/B978-0-444-53860-4.00010-6. Prog Brain Res. 2012. PMID: 22230629 Review.
-
Noether's symmetry and conserved quantity for a time-delayed Hamiltonian system of Herglotz type.R Soc Open Sci. 2018 Oct 3;5(10):180208. doi: 10.1098/rsos.180208. eCollection 2018 Oct. R Soc Open Sci. 2018. PMID: 30473803 Free PMC article.
-
Disruption in the inhibitory architecture of the cell minicolumn: implications for autism.Neuroscientist. 2003 Dec;9(6):496-507. doi: 10.1177/1073858403253552. Neuroscientist. 2003. PMID: 14678582 Review.
Cited by
-
Predicting age from cortical structure across the lifespan.Eur J Neurosci. 2018 Mar;47(5):399-416. doi: 10.1111/ejn.13835. Epub 2018 Feb 12. Eur J Neurosci. 2018. PMID: 29359873 Free PMC article.
-
A computational model for the loss of neuronal organization in microcolumns.Biophys J. 2014 May 20;106(10):2233-42. doi: 10.1016/j.bpj.2014.04.012. Biophys J. 2014. PMID: 24853752 Free PMC article.
-
Inter-laminar microcircuits across neocortex: repair and augmentation.Front Syst Neurosci. 2013 Nov 19;7:80. doi: 10.3389/fnsys.2013.00080. eCollection 2013. Front Syst Neurosci. 2013. PMID: 24312019 Free PMC article. No abstract available.
-
Prefrontal cortical minicolumn: from executive control to disrupted cognitive processing.Brain. 2014 Jul;137(Pt 7):1863-75. doi: 10.1093/brain/awt359. Epub 2014 Feb 14. Brain. 2014. PMID: 24531625 Free PMC article. Review.
-
Three Decades of Valproate: A Current Model for Studying Autism Spectrum Disorder.Curr Neuropharmacol. 2024;22(2):260-289. doi: 10.2174/1570159X22666231003121513. Curr Neuropharmacol. 2024. PMID: 37873949 Free PMC article. Review.
References
-
- Amundson R. (1994). Two concepts of constraint: adaptationism and the challenge from developmental biology. Philos. Sci. 61, 556–57810.1086/289822 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources