

Assembly of Drosophila centromeric nucleosomes requires CID dimerization
- PMID: 22209075
- PMCID: PMC3299411
- DOI: 10.1016/j.molcel.2011.12.010
Assembly of Drosophila centromeric nucleosomes requires CID dimerization
Abstract
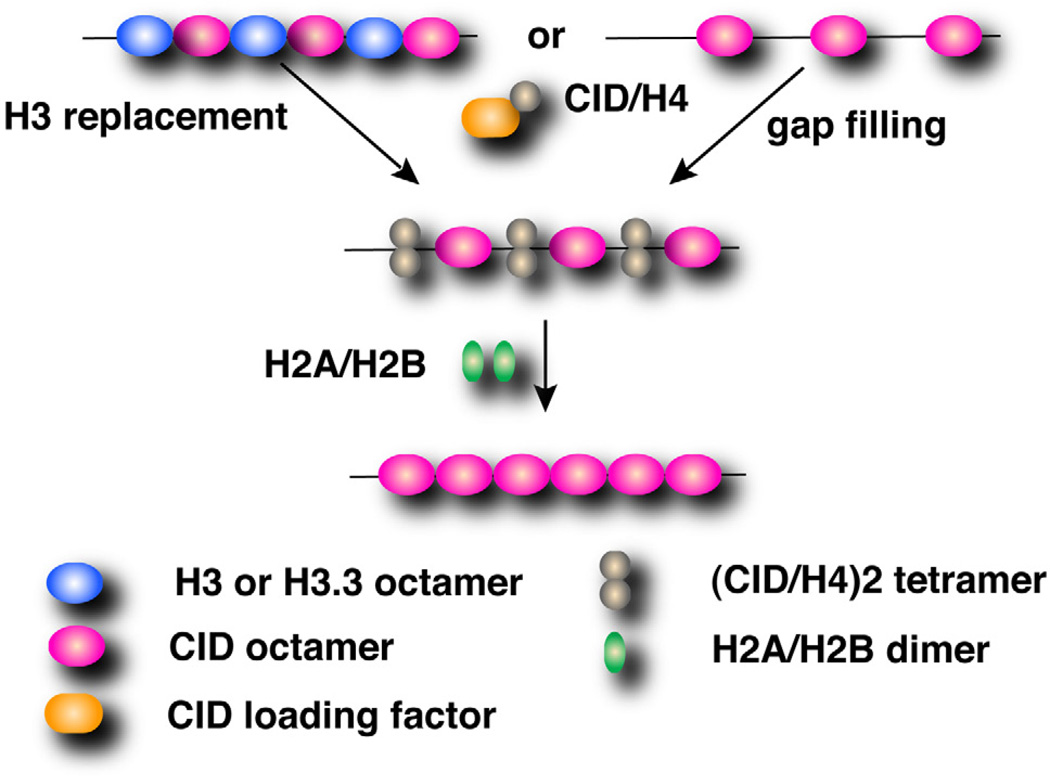
Centromeres are essential chromosomal regions required for kinetochore assembly and chromosome segregation. The composition and organization of centromeric nucleosomes containing the essential histone H3 variant CENP-A (CID in Drosophila) is a fundamental, unresolved issue. Using immunoprecipitation of CID mononucleosomes and cysteine crosslinking, we demonstrate that centromeric nucleosomes contain CID dimers in vivo. Furthermore, CID dimerization and centromeric targeting require a residue implicated in formation of the four-helix bundle, which mediates intranucleosomal H3 dimerization and nucleosome integrity. Taken together, our findings suggest that CID nucleosomes are octameric in vivo and that CID dimerization is essential for correct centromere assembly.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
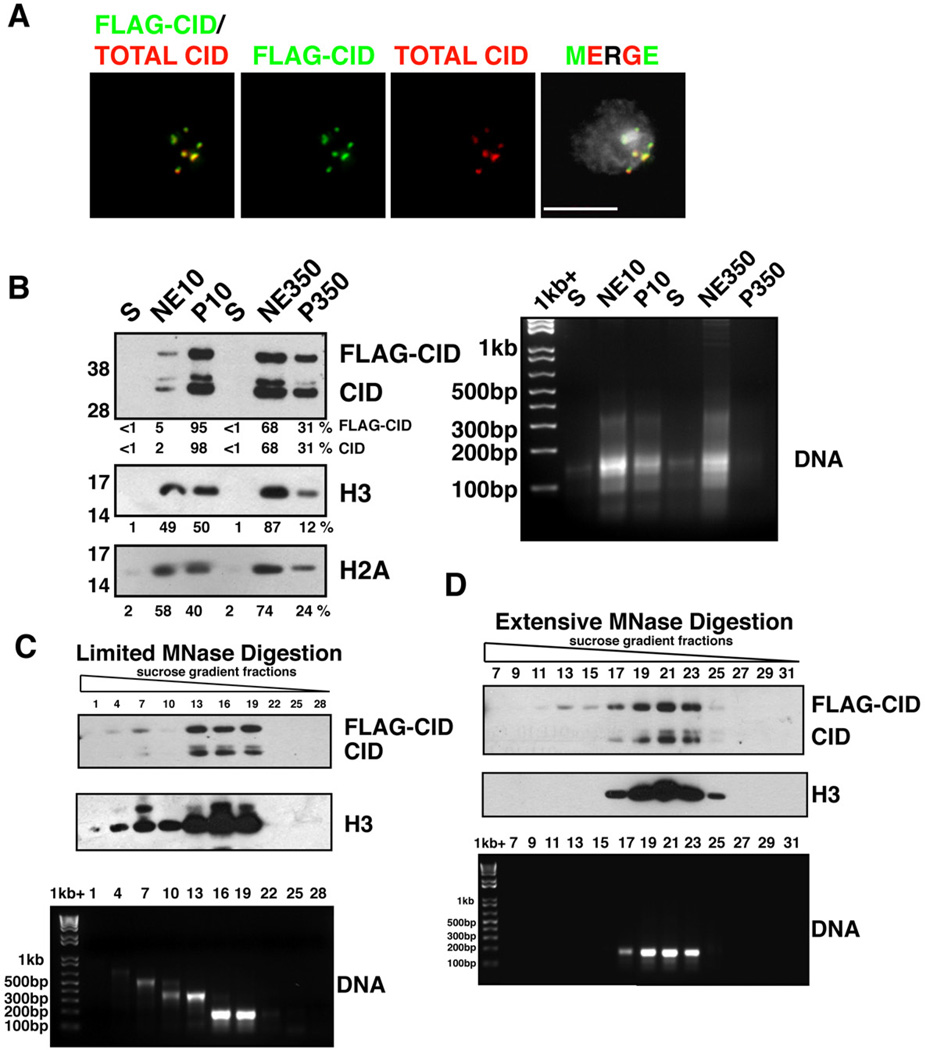
FLAG-CID (green) and total CID (red) colocalize at centromeres in a stable S2 cell line. FLAG-CID and total CID were stained by α-FLAG and α-CID antibodies, respectively. DNA staining by DAPI (grey). Scale bar = 5 µm.
(left) FLAG-CID and endogenous CID are more resistant to salt extraction than H3. After MNase digestion, soluble nucleoplasmic fractions (S), chromatin extracted with either 10 mM or 350 mM salt (NE10 and NE350, respectively), and insoluble nuclear pellets (P10 and P350, respectively) were analyzed by Western blots for CID, H3 and H2A. (right) DNA agarose gel with the same samples.
(top) FLAG-CID and endogenous CID co-fractionate with peaks of H3 in sucrose gradient fractions after limited MNase digestion. (bottom) DNA agarose gel shows sizes consistent with mainly mono-, di- or tri-nucleosomes.
(Top) FLAG-CID and endogenous CID co-migrate with H3 in sucrose gradient fractions after extensive MNase digestion. (bottom) DNA agarose gel from the same samples.
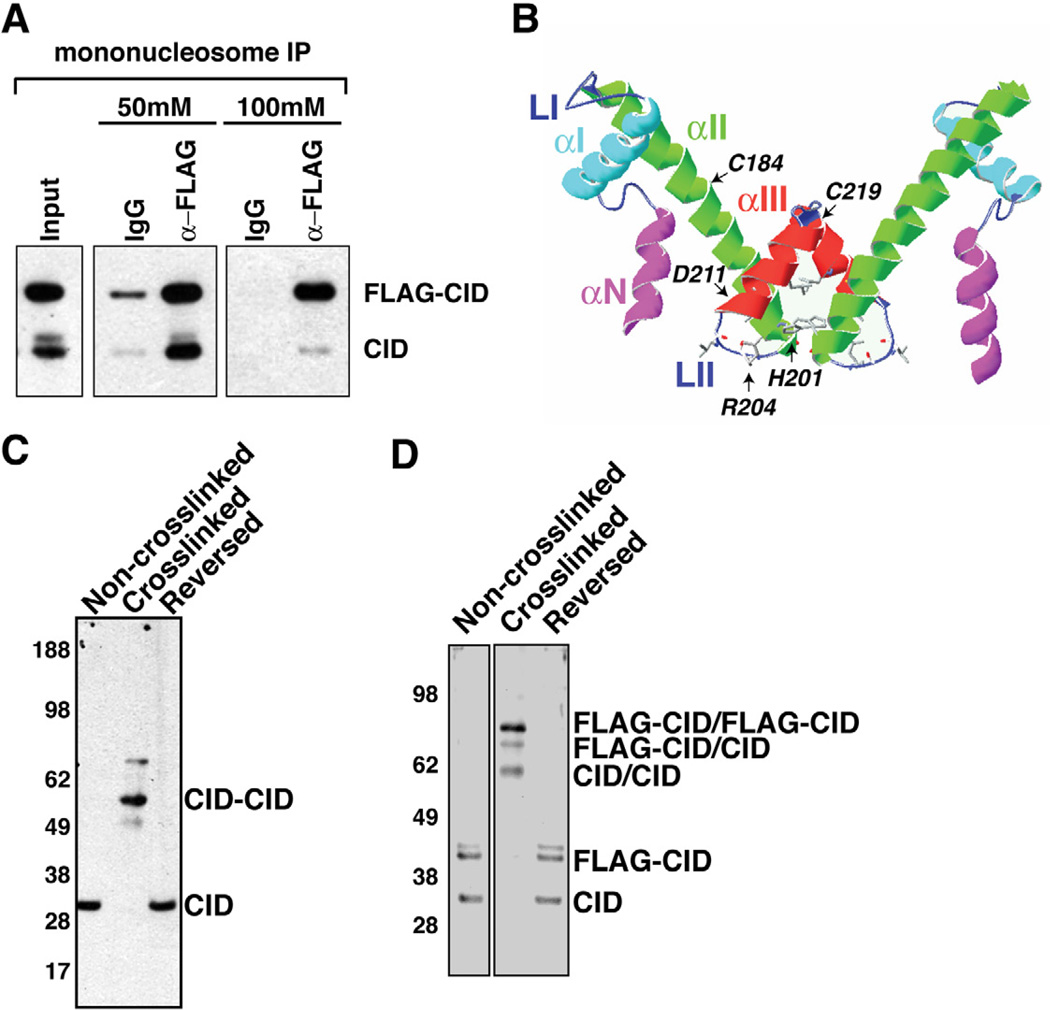
Endogenous CID coimmunoprecipitates with FLAG-CID from mononucleosomes. Inputs consist of mononucleosomes. α-mouse IgG beads were used in control IPs (IgG).
Schematic display of the HFD of CID using a Cse4 model as reference (Bloom et al., 2006). Helices- αN, -αI, -αII, -αIII, and Loops LI, LII are indicated. CATD is comprised of LI and αII. The putative four helix bundle is comprised of the C terminal halves of αII and αIII.
Cysteine crosslinking of S2 chromatin. CID monomers migrate at 30 kDa without crosslinking and after crosslink reversal. After crosslinking, the majority of CID migrate at 60 kDa, consistent with the size of dimers.
Cysteine crosslinking of FLAG-CID chromatin. FLAG-CID and endogenous CID migrate as monomers before crosslinking and after crosslink reversal. After crosslinking, three major bands were detected by CID antibody that correspond to CID/CID, FLAG-CID/CID and FLAG-CID/FLAG-CID, respectively.
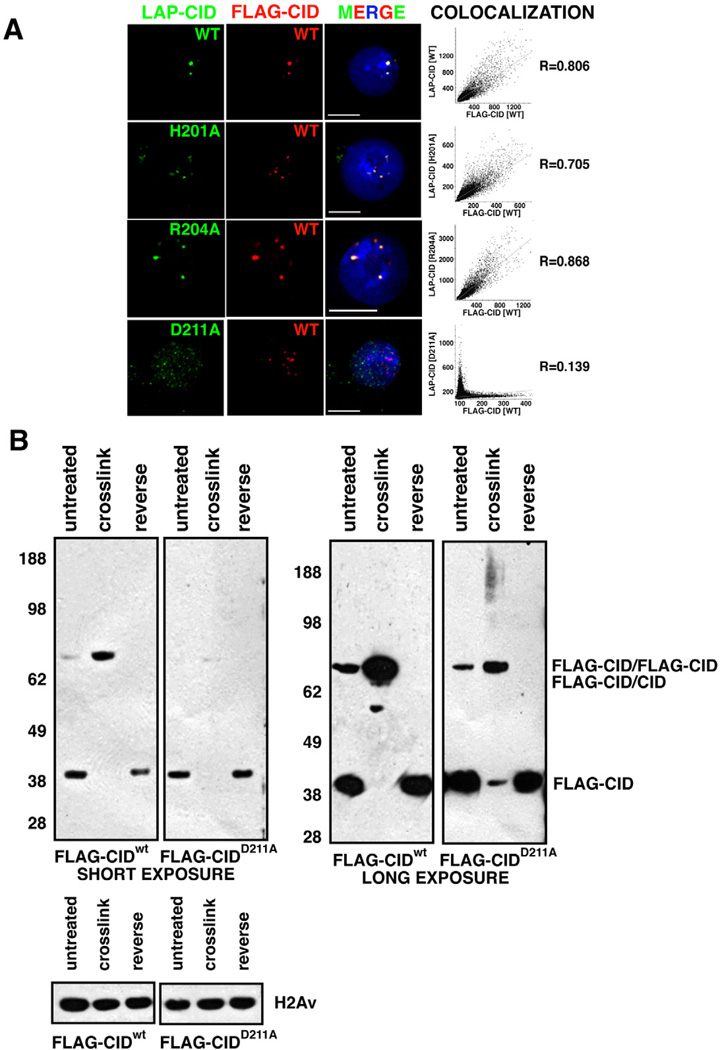
Localization of mutant LAP-CID constructs. FLAG-CIDwt (red) at centromeres colocalizes with LAP-CIDwt (98.5%, n=130)(green), and point mutants LAP-CIDH201A (99.0%, n=96) (green) and LAP-CIDR204A (97.3%, n=112) (green). In contrast, few LAP-CIDD211A (2.63%, n=152) (green) colocalize with FLAG-CIDwt (red), and often associates with non-centromeric regions. Scale bars = 5 µm. The graphs show Pearson’s coefficient of correlation (R) of the individual images.
FLAG-CIDD211A mutant proteins are defective for dimerization. FLAG-CIDwt was efficiently crosslinked as dimers. In contrast, FLAG-CIDD211A crosslinking converted ~43% into dimmers (short exposure) and 40% into a smear without distinct bands (long exposure), while 17% remained as monomers. Total CIDD211A intensity after crosslinking was calculated as the sum of monomers, dimers and smear. H2Av, which lacks cysteines, was used as loading control.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
