

Arabidopsis VILLIN2 and VILLIN3 are required for the generation of thick actin filament bundles and for directional organ growth
- PMID: 22209875
- PMCID: PMC3291277
- DOI: 10.1104/pp.111.192385
Arabidopsis VILLIN2 and VILLIN3 are required for the generation of thick actin filament bundles and for directional organ growth
Abstract
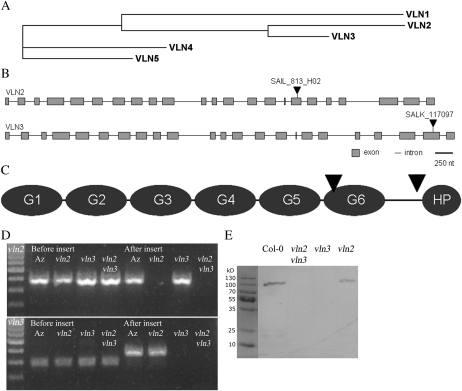
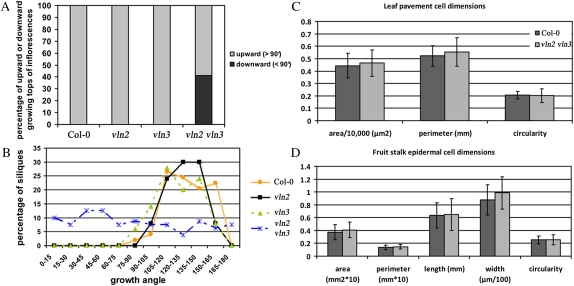
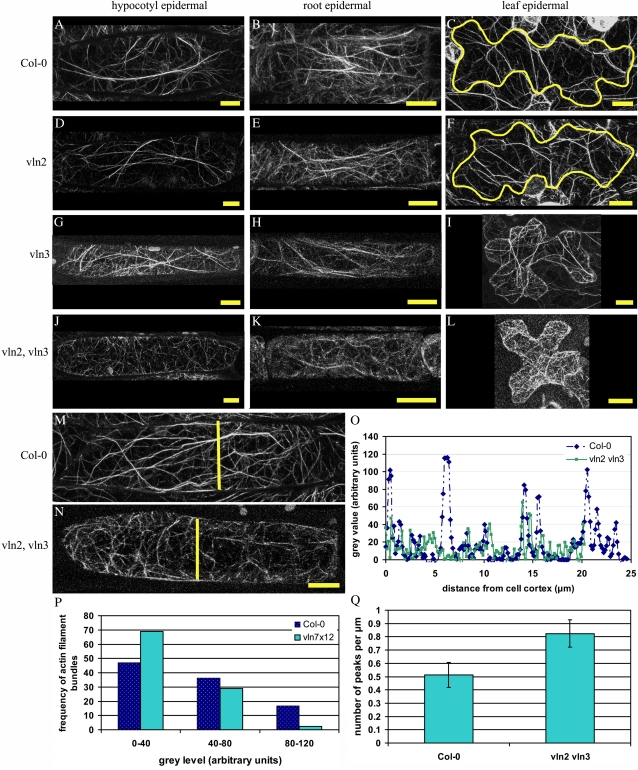
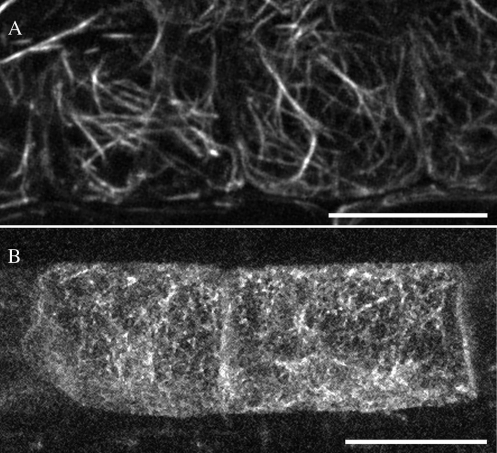
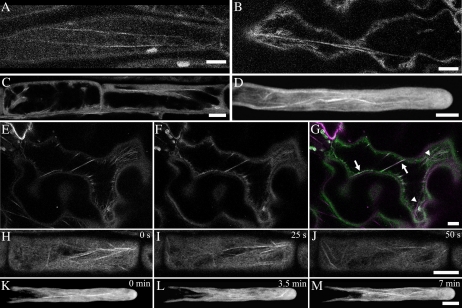
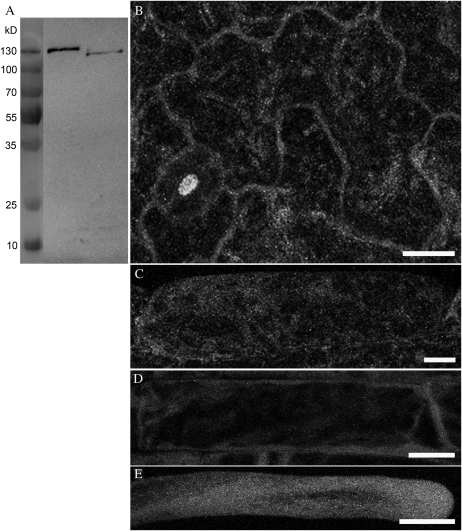
In plant cells, actin filament bundles serve as tracks for myosin-dependent organelle movement and play a role in the organization of the cytoplasm. Although virtually all plant cells contain actin filament bundles, the role of the different actin-bundling proteins remains largely unknown. In this study, we investigated the role of the actin-bundling protein villin in Arabidopsis (Arabidopsis thaliana). We used Arabidopsis T-DNA insertion lines to generate a double mutant in which VILLIN2 (VLN2) and VLN3 transcripts are truncated. Leaves, stems, siliques, and roots of vln2 vln3 double mutant plants are twisted, which is caused by local differences in cell length. Microscopy analysis of the actin cytoskeleton showed that in these double mutant plants, thin actin filament bundles are more abundant while thick actin filament bundles are virtually absent. In contrast to full-length VLN3, truncated VLN3 lacking the headpiece region does not rescue the phenotype of the vln2 vln3 double mutant. Our results show that villin is involved in the generation of thick actin filament bundles in several cell types and suggest that these bundles are involved in the regulation of coordinated cell expansion.
Figures

Similar articles
-
Arabidopsis VILLIN2 and VILLIN3 act redundantly in sclerenchyma development via bundling of actin filaments.Plant J. 2012 Sep;71(6):962-75. doi: 10.1111/j.1365-313X.2012.05044.x. Epub 2012 Jun 28. Plant J. 2012. PMID: 22563899
-
Arabidopsis villins promote actin turnover at pollen tube tips and facilitate the construction of actin collars.Plant Cell. 2013 May;25(5):1803-17. doi: 10.1105/tpc.113.110940. Epub 2013 May 28. Plant Cell. 2013. PMID: 23715472 Free PMC article.
-
GhVLN4 is involved in cell elongation via regulation of actin organization.Planta. 2017 Oct;246(4):687-700. doi: 10.1007/s00425-017-2723-7. Epub 2017 Jun 24. Planta. 2017. PMID: 28647813
-
Actin-myosin XI: an intracellular control network in plants.Biochem Biophys Res Commun. 2018 Nov 25;506(2):403-408. doi: 10.1016/j.bbrc.2017.12.169. Epub 2018 Jan 5. Biochem Biophys Res Commun. 2018. PMID: 29307817 Review.
-
Dynamic behavior of plastids related to environmental response.Curr Opin Plant Biol. 2012 Dec;15(6):722-8. doi: 10.1016/j.pbi.2012.08.003. Epub 2012 Aug 29. Curr Opin Plant Biol. 2012. PMID: 22939249 Review.
Cited by
-
Streamlined spatial and environmental expression signatures characterize the minimalist duckweed Wolffia australiana.Genome Res. 2024 Aug 20;34(7):1106-1120. doi: 10.1101/gr.279091.124. Genome Res. 2024. PMID: 38951025 Free PMC article.
-
Morphometric analysis of actin networks.Mol Biol Cell. 2024 Dec 1;35(12):ar146. doi: 10.1091/mbc.E24-06-0248. Epub 2024 Oct 23. Mol Biol Cell. 2024. PMID: 39441713 Free PMC article.
-
Functional Characterization of PeVLN4 Involved in Regulating Pollen Tube Growth from Passion Fruit.Int J Mol Sci. 2025 Mar 6;26(5):2348. doi: 10.3390/ijms26052348. Int J Mol Sci. 2025. PMID: 40076967 Free PMC article.
-
Bundling actin filaments from membranes: some novel players.Front Plant Sci. 2012 Aug 24;3:188. doi: 10.3389/fpls.2012.00188. eCollection 2012. Front Plant Sci. 2012. PMID: 22936939 Free PMC article.
-
VILLIN2 regulates cotton defense against Verticillium dahliae by modulating actin cytoskeleton remodeling.Plant Physiol. 2023 May 2;192(1):666-679. doi: 10.1093/plphys/kiad095. Plant Physiol. 2023. PMID: 36881883 Free PMC article.
References
-
- Bretscher A, Weber K. (1980) Villin is a major protein of the microvillus cytoskeleton which binds both G and F actin in a calcium-dependent manner. Cell 20: 839–847 - PubMed
-
- Bryan J, Kurth MC. (1984) Actin-gelsolin interactions: evidence for two actin-binding sites. J Biol Chem 259: 7480–7487 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
