

The response to heat shock and oxidative stress in Saccharomyces cerevisiae
- PMID: 22209905
- PMCID: PMC3316637
- DOI: 10.1534/genetics.111.128033
The response to heat shock and oxidative stress in Saccharomyces cerevisiae
Abstract
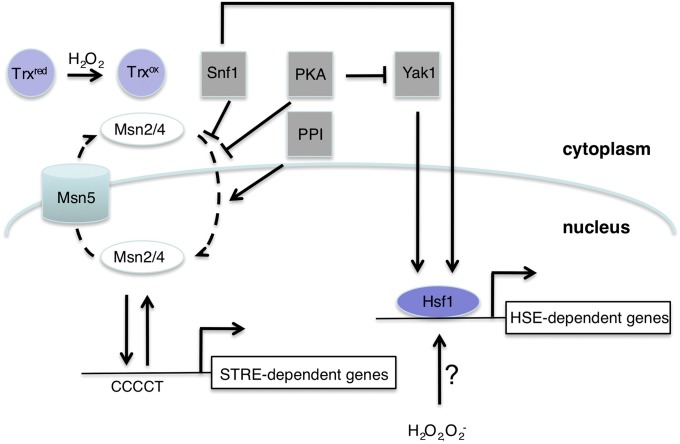
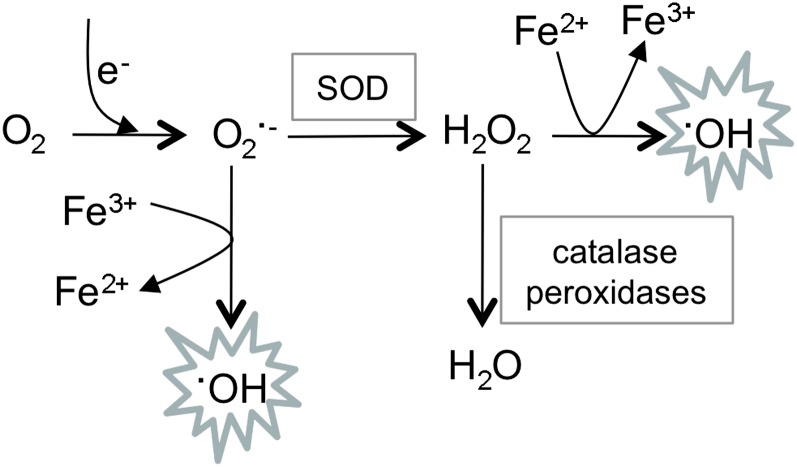
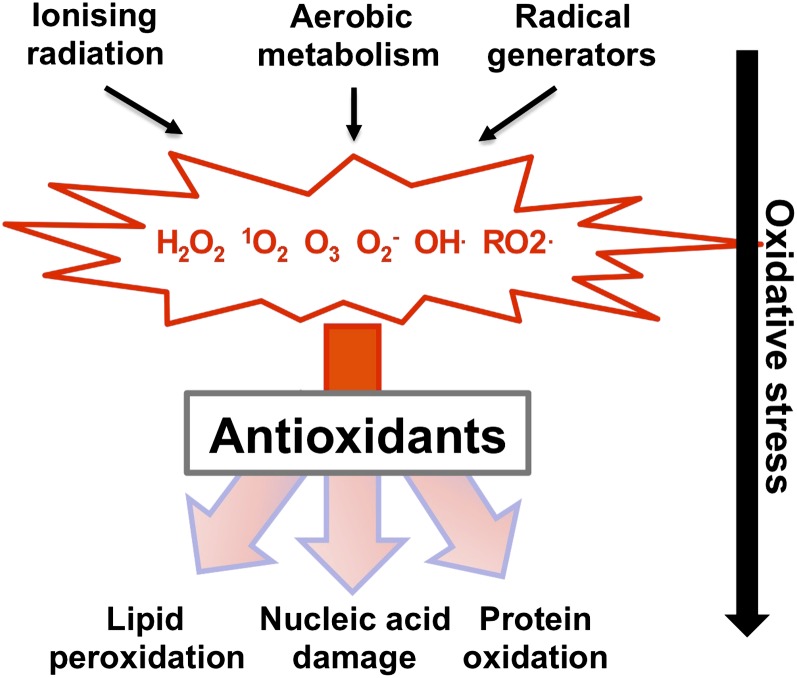
A common need for microbial cells is the ability to respond to potentially toxic environmental insults. Here we review the progress in understanding the response of the yeast Saccharomyces cerevisiae to two important environmental stresses: heat shock and oxidative stress. Both of these stresses are fundamental challenges that microbes of all types will experience. The study of these environmental stress responses in S. cerevisiae has illuminated many of the features now viewed as central to our understanding of eukaryotic cell biology. Transcriptional activation plays an important role in driving the multifaceted reaction to elevated temperature and levels of reactive oxygen species. Advances provided by the development of whole genome analyses have led to an appreciation of the global reorganization of gene expression and its integration between different stress regimens. While the precise nature of the signal eliciting the heat shock response remains elusive, recent progress in the understanding of induction of the oxidative stress response is summarized here. Although these stress conditions represent ancient challenges to S. cerevisiae and other microbes, much remains to be learned about the mechanisms dedicated to dealing with these environmental parameters.
Figures

Similar articles
-
The Skn7 response regulator of Saccharomyces cerevisiae interacts with Hsf1 in vivo and is required for the induction of heat shock genes by oxidative stress.Mol Biol Cell. 2000 Jul;11(7):2335-47. doi: 10.1091/mbc.11.7.2335. Mol Biol Cell. 2000. PMID: 10888672 Free PMC article.
-
Regulation of thermotolerance by stress-induced transcription factors in Saccharomyces cerevisiae.Eukaryot Cell. 2008 May;7(5):783-90. doi: 10.1128/EC.00029-08. Epub 2008 Mar 21. Eukaryot Cell. 2008. PMID: 18359875 Free PMC article.
-
A stress regulatory network for co-ordinated activation of proteasome expression mediated by yeast heat shock transcription factor.Mol Microbiol. 2006 Apr;60(1):240-51. doi: 10.1111/j.1365-2958.2006.05097.x. Mol Microbiol. 2006. PMID: 16556235
-
Regulation of the heat shock transcription factor Hsf1 in fungi: implications for temperature-dependent virulence traits.FEMS Yeast Res. 2018 Aug 1;18(5):foy041. doi: 10.1093/femsyr/foy041. FEMS Yeast Res. 2018. PMID: 29788061 Free PMC article. Review.
-
[Alternative ways of stress regulation in cells of Saccharomyces cerevisiae: transcriptional activators Msn2 and Msn4].Tsitologiia. 2009;51(3):271-8. Tsitologiia. 2009. PMID: 19435282 Review. Russian.
Cited by
-
Heat Stress Modulates Mycelium Growth, Heat Shock Protein Expression, Ganoderic Acid Biosynthesis, and Hyphal Branching of Ganoderma lucidum via Cytosolic Ca2.Appl Environ Microbiol. 2016 Jun 30;82(14):4112-4125. doi: 10.1128/AEM.01036-16. Print 2016 Jul 15. Appl Environ Microbiol. 2016. PMID: 27129961 Free PMC article.
-
Understanding the Mechanism of Thermotolerance Distinct From Heat Shock Response Through Proteomic Analysis of Industrial Strains of Saccharomyces cerevisiae.Mol Cell Proteomics. 2015 Jul;14(7):1885-97. doi: 10.1074/mcp.M114.045781. Epub 2015 Apr 29. Mol Cell Proteomics. 2015. PMID: 25926660 Free PMC article.
-
Bioavailability and Bioactivities of Polyphenols Eco Extracts from Coffee Grounds after In Vitro Digestion.Foods. 2020 Sep 12;9(9):1281. doi: 10.3390/foods9091281. Foods. 2020. PMID: 32932599 Free PMC article.
-
Mitochondria-Associated Degradation Pathway (MAD) Function beyond the Outer Membrane.Cell Rep. 2020 Jul 14;32(2):107902. doi: 10.1016/j.celrep.2020.107902. Cell Rep. 2020. PMID: 32668258 Free PMC article.
-
The InsP7 phosphatase Siw14 regulates inositol pyrophosphate levels to control localization of the general stress response transcription factor Msn2.J Biol Chem. 2020 Feb 14;295(7):2043-2056. doi: 10.1074/jbc.RA119.012148. Epub 2019 Dec 17. J Biol Chem. 2020. PMID: 31848224 Free PMC article.
References
-
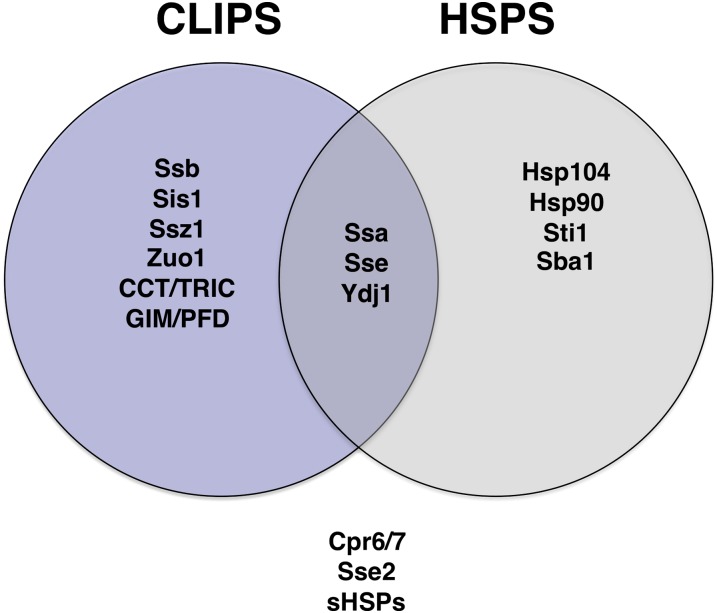
- Albanese V., Yam A. Y., Baughman J., Parnot C., Frydman J., 2006. Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell 124: 75–88 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
