

Acute depletion of Tet1-dependent 5-hydroxymethylcytosine levels impairs LIF/Stat3 signaling and results in loss of embryonic stem cell identity
- PMID: 22210859
- PMCID: PMC3333871
- DOI: 10.1093/nar/gkr1253
Acute depletion of Tet1-dependent 5-hydroxymethylcytosine levels impairs LIF/Stat3 signaling and results in loss of embryonic stem cell identity
Abstract
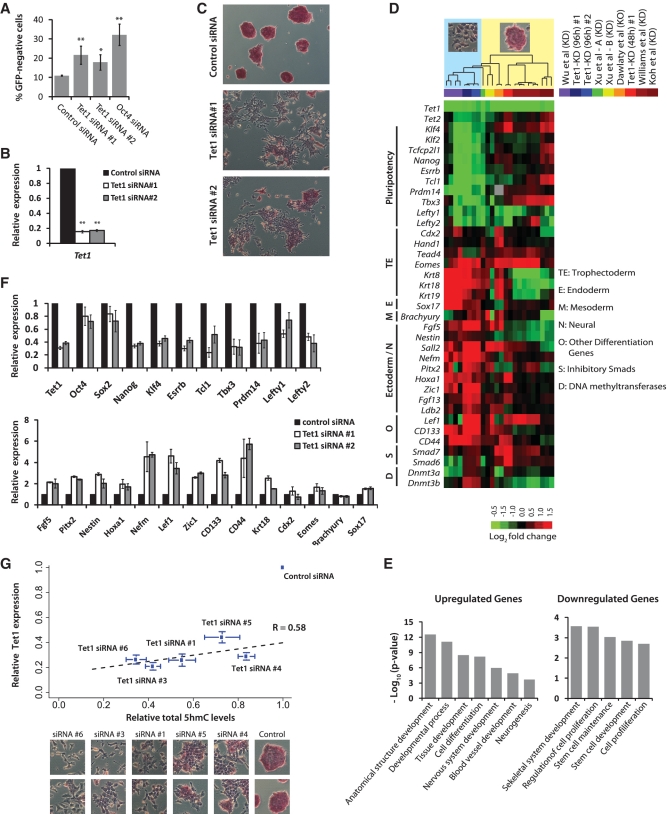
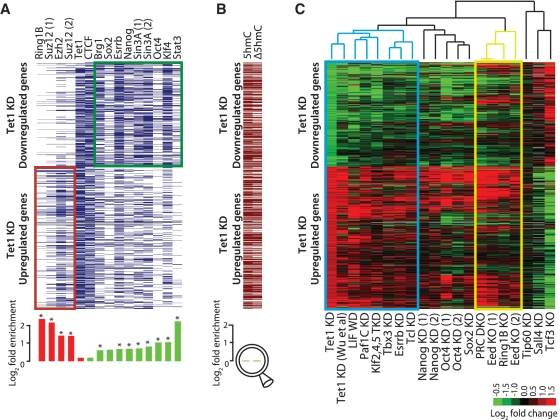
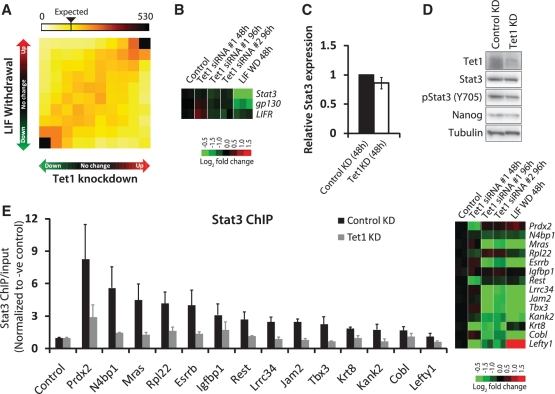
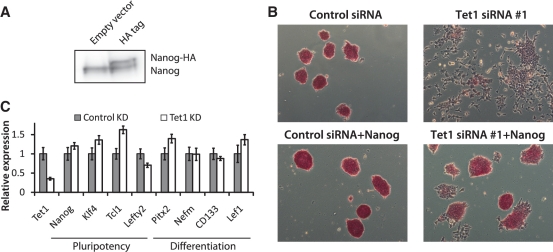
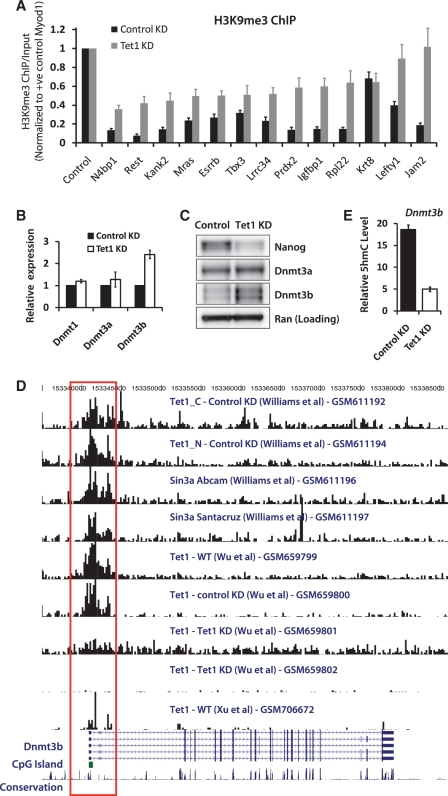
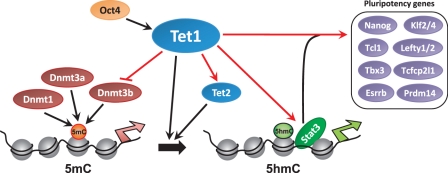
The TET family of FE(II) and 2-oxoglutarate-dependent enzymes (Tet1/2/3) promote DNA demethylation by converting 5-methylcytosine to 5-hydroxymethylcytosine (5hmC), which they further oxidize into 5-formylcytosine and 5-carboxylcytosine. Tet1 is robustly expressed in mouse embryonic stem cells (mESCs) and has been implicated in mESC maintenance. Here we demonstrate that, unlike genetic deletion, RNAi-mediated depletion of Tet1 in mESCs led to a significant reduction in 5hmC and loss of mESC identity. The differentiation phenotype due to Tet1 depletion positively correlated with the extent of 5hmC loss. Meta-analyses of genomic data sets suggested interaction between Tet1 and leukemia inhibitory factor (LIF) signaling. LIF signaling is known to promote self-renewal and pluripotency in mESCs partly by opposing MAPK/ERK-mediated differentiation. Withdrawal of LIF leads to differentiation of mESCs. We discovered that Tet1 depletion impaired LIF-dependent Stat3-mediated gene activation by affecting Stat3's ability to bind to its target sites on chromatin. Nanog overexpression or inhibition of MAPK/ERK signaling, both known to maintain mESCs in the absence of LIF, rescued Tet1 depletion, further supporting the dependence of LIF/Stat3 signaling on Tet1. These data support the conclusion that analysis of mESCs in the hours/days immediately following efficient Tet1 depletion reveals Tet1's normal physiological role in maintaining the pluripotent state that may be subject to homeostatic compensation in genetic models.
Figures
Similar articles
-
Genome-wide regulation of 5hmC, 5mC, and gene expression by Tet1 hydroxylase in mouse embryonic stem cells.Mol Cell. 2011 May 20;42(4):451-64. doi: 10.1016/j.molcel.2011.04.005. Epub 2011 Apr 21. Mol Cell. 2011. PMID: 21514197 Free PMC article.
-
Foxm1 mediates LIF/Stat3-dependent self-renewal in mouse embryonic stem cells and is essential for the generation of induced pluripotent stem cells.PLoS One. 2014 Apr 17;9(4):e92304. doi: 10.1371/journal.pone.0092304. eCollection 2014. PLoS One. 2014. Retraction in: PLoS One. 2019 Jul 5;14(7):e0219580. doi: 10.1371/journal.pone.0219580. PMID: 24743237 Free PMC article. Retracted.
-
Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification.Nature. 2010 Aug 26;466(7310):1129-33. doi: 10.1038/nature09303. Nature. 2010. PMID: 20639862 Free PMC article.
-
Mechanisms and functions of Tet protein-mediated 5-methylcytosine oxidation.Genes Dev. 2011 Dec 1;25(23):2436-52. doi: 10.1101/gad.179184.111. Genes Dev. 2011. PMID: 22156206 Free PMC article. Review.
-
Regulation of embryonic stem cell self-renewal and pluripotency by leukaemia inhibitory factor.Biochem J. 2011 Aug 15;438(1):11-23. doi: 10.1042/BJ20102152. Biochem J. 2011. PMID: 21793804 Free PMC article. Review.
Cited by
-
What do DNA methylation studies tell us about depression? A systematic review.Transl Psychiatry. 2019 Feb 4;9(1):68. doi: 10.1038/s41398-019-0412-y. Transl Psychiatry. 2019. PMID: 30718449 Free PMC article.
-
Distinct and overlapping control of 5-methylcytosine and 5-hydroxymethylcytosine by the TET proteins in human cancer cells.Genome Biol. 2014 Jun 23;15(6):R81. doi: 10.1186/gb-2014-15-6-r81. Genome Biol. 2014. PMID: 24958354 Free PMC article.
-
Leukemia Inhibitory Factor Promotes Aggressiveness of Chordoma.Oncol Res. 2017 Aug 7;25(7):1177-1188. doi: 10.3727/096504017X14874349473815. Epub 2017 Feb 28. Oncol Res. 2017. PMID: 28247842 Free PMC article.
-
Tet1-dependent epigenetic modification of BDNF expression in dorsal horn neurons mediates neuropathic pain in rats.Sci Rep. 2016 Nov 18;6:37411. doi: 10.1038/srep37411. Sci Rep. 2016. PMID: 27857218 Free PMC article.
-
Integrative framework for identification of key cell identity genes uncovers determinants of ES cell identity and homeostasis.Proc Natl Acad Sci U S A. 2014 Apr 22;111(16):E1581-90. doi: 10.1073/pnas.1318598111. Epub 2014 Apr 7. Proc Natl Acad Sci U S A. 2014. PMID: 24711389 Free PMC article.
References
-
- Nichols J, Smith A. Naive and primed pluripotent states. Cell Stem Cell. 2009;4:487–492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
