

Targeting self- and foreign antigens to dendritic cells via DC-ASGPR generates IL-10-producing suppressive CD4+ T cells
- PMID: 22213806
- PMCID: PMC3260876
- DOI: 10.1084/jem.20110399
Targeting self- and foreign antigens to dendritic cells via DC-ASGPR generates IL-10-producing suppressive CD4+ T cells
Abstract
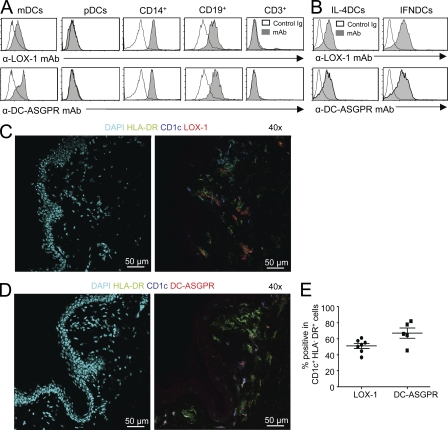
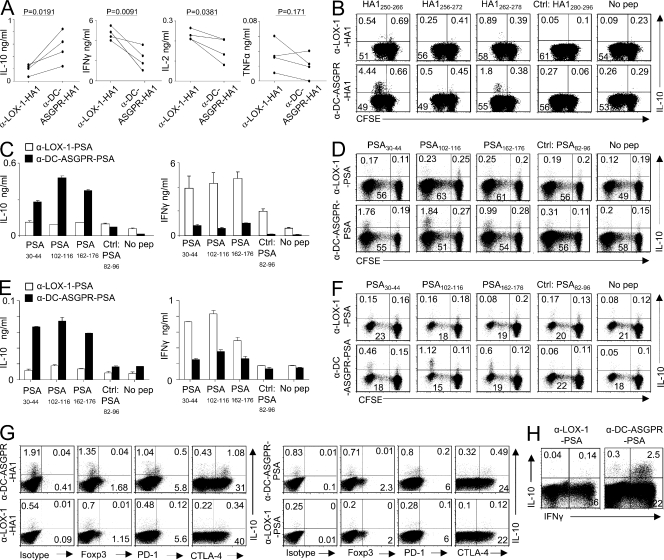
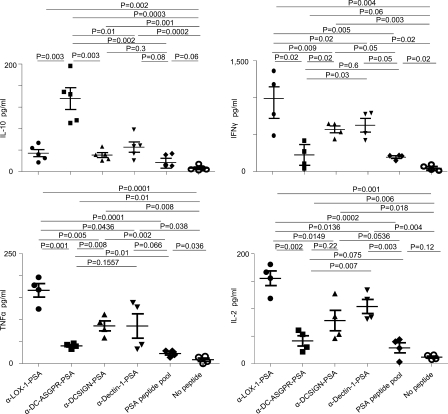
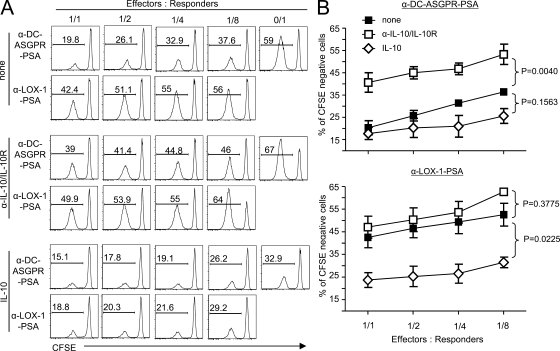
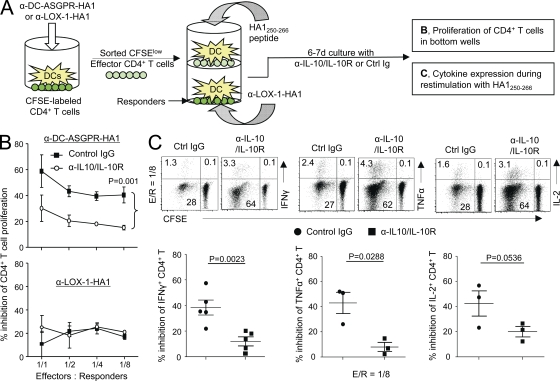
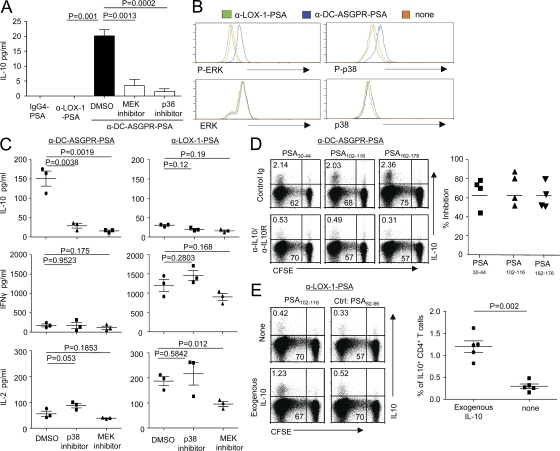
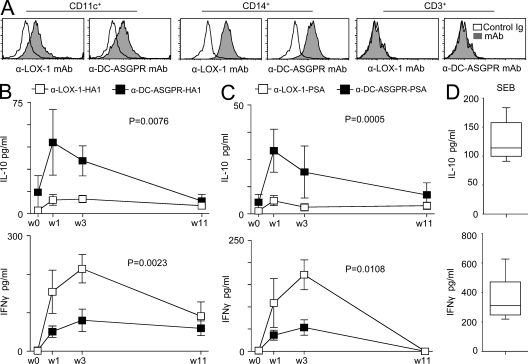
Dendritic cells (DCs) can initiate and shape host immune responses toward either immunity or tolerance by their effects on antigen-specific CD4(+) T cells. DC-asialoglycoprotein receptor (DC-ASGPR), a lectinlike receptor, is a known scavenger receptor. Here, we report that targeting antigens to human DCs via DC-ASGPR, but not lectin-like oxidized-LDL receptor, Dectin-1, or DC-specific ICAM-3-grabbing nonintegrin favors the generation of antigen-specific suppressive CD4(+) T cells that produce interleukin 10 (IL-10). These findings apply to both self- and foreign antigens, as well as memory and naive CD4(+) T cells. The generation of such IL-10-producing CD4(+) T cells requires p38/extracellular signal-regulated kinase phosphorylation and IL-10 induction in DCs. We further demonstrate that immunization of nonhuman primates with antigens fused to anti-DC-ASGPR monoclonal antibody generates antigen-specific CD4(+) T cells that produce IL-10 in vivo. This study provides a new strategy for the establishment of antigen-specific IL-10-producing suppressive T cells in vivo by targeting whole protein antigens to DCs via DC-ASGPR.
Figures
References
-
- Bacchetta R., Bigler M., Touraine J.L., Parkman R., Tovo P.A., Abrams J., de Waal Malefyt R., de Vries J.E., Roncarolo M.G. 1994. High levels of interleukin 10 production in vivo are associated with tolerance in SCID patients transplanted with HLA mismatched hematopoietic stem cells. J. Exp. Med. 179:493–502 10.1084/jem.179.2.493 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
