

MpeR regulates the mtr efflux locus in Neisseria gonorrhoeae and modulates antimicrobial resistance by an iron-responsive mechanism
- PMID: 22214775
- PMCID: PMC3294918
- DOI: 10.1128/AAC.06112-11
MpeR regulates the mtr efflux locus in Neisseria gonorrhoeae and modulates antimicrobial resistance by an iron-responsive mechanism
Abstract
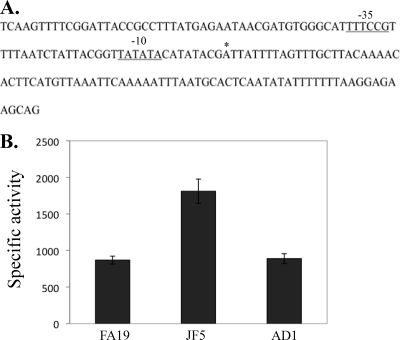
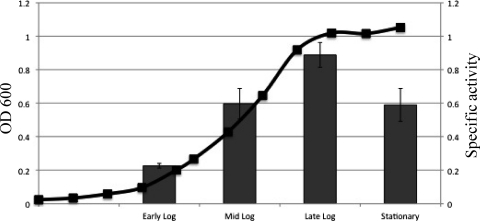
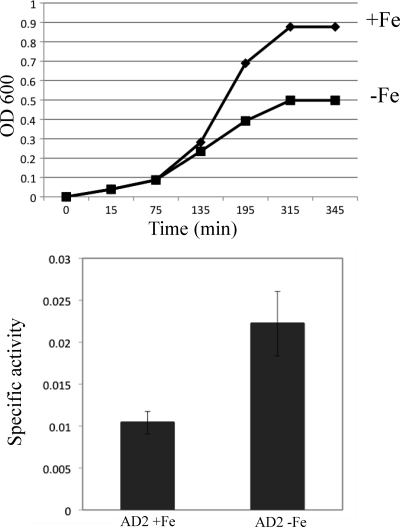
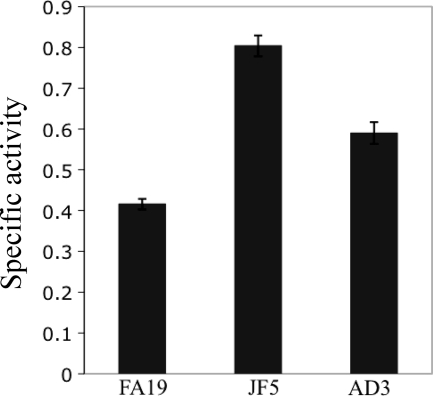
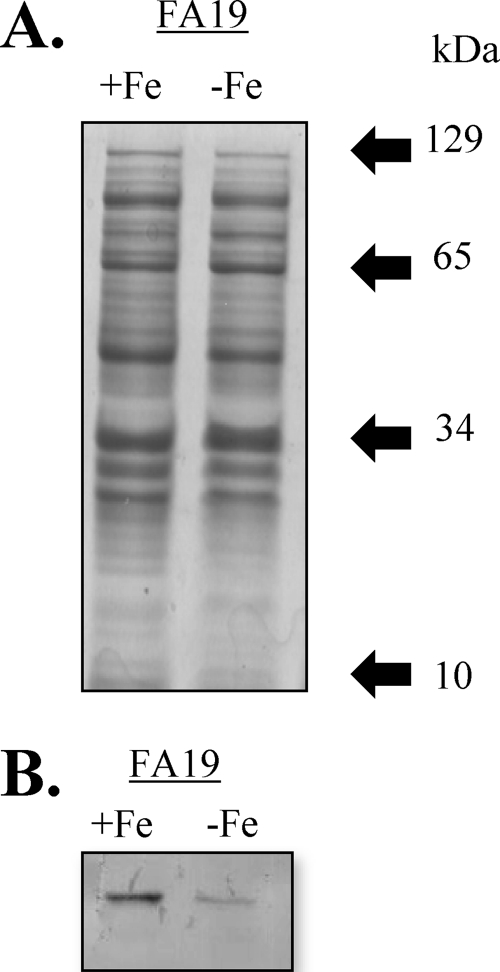
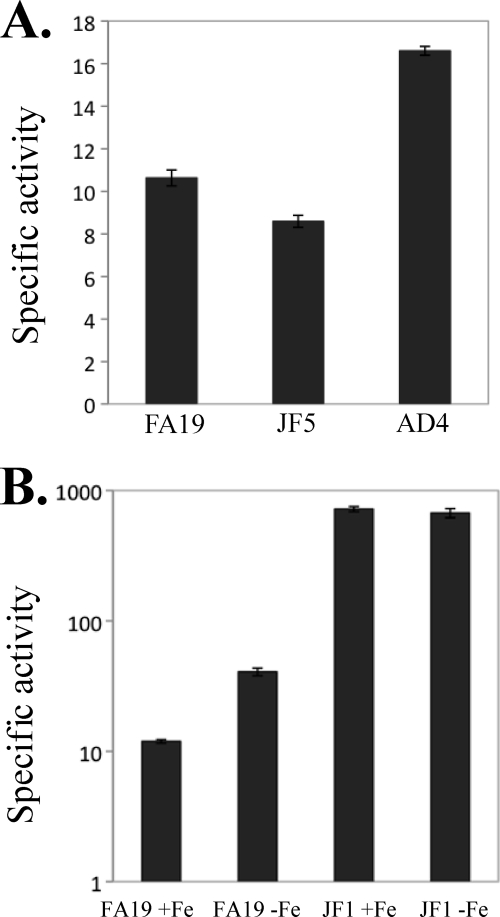
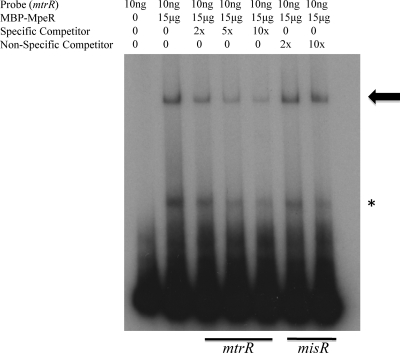
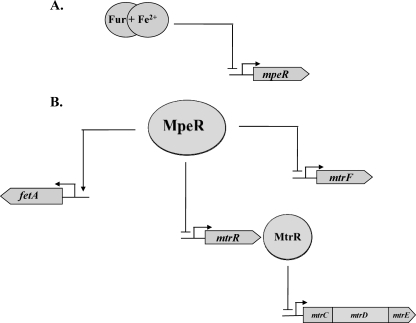
Previous studies have shown that the MpeR transcriptional regulator produced by Neisseria gonorrhoeae represses the expression of mtrF, which encodes a putative inner membrane protein (MtrF). MtrF works as an accessory protein with the Mtr efflux pump, helping gonococci to resist high levels of diverse hydrophobic antimicrobials. Regulation of mpeR has been reported to occur by an iron-dependent mechanism involving Fur (ferric uptake regulator). Collectively, these observations suggest the presence of an interconnected regulatory system in gonococci that modulates the expression of efflux pump protein-encoding genes in an iron-responsive manner. Herein, we describe this connection and report that levels of gonococcal resistance to a substrate of the mtrCDE-encoded efflux pump can be modulated by MpeR and the availability of free iron. Using microarray analysis, we found that the mtrR gene, which encodes a direct repressor (MtrR) of mtrCDE, is an MpeR-repressed determinant in the late logarithmic phase of growth when free iron levels would be reduced due to bacterial consumption. This repression was enhanced under conditions of iron limitation and resulted in increased expression of the mtrCDE efflux pump operon. Furthermore, as judged by DNA-binding analysis, MpeR-mediated repression of mtrR was direct. Collectively, our results indicate that both genetic and physiologic parameters (e.g., iron availability) can influence the expression of the mtr efflux system and modulate levels of gonococcal susceptibility to efflux pump substrates.
Figures
Similar articles
-
Regulation of mtrF expression in Neisseria gonorrhoeae and its role in high-level antimicrobial resistance.J Bacteriol. 2005 Jun;187(11):3713-20. doi: 10.1128/JB.187.11.3713-3720.2005. J Bacteriol. 2005. PMID: 15901695 Free PMC article.
-
Could Dampening Expression of the Neisseria gonorrhoeae mtrCDE-Encoded Efflux Pump Be a Strategy To Preserve Currently or Resurrect Formerly Used Antibiotics To Treat Gonorrhea?mBio. 2019 Aug 13;10(4):e01576-19. doi: 10.1128/mBio.01576-19. mBio. 2019. PMID: 31409679 Free PMC article.
-
Identification of a cell envelope protein (MtrF) involved in hydrophobic antimicrobial resistance in Neisseria gonorrhoeae.J Antimicrob Chemother. 2003 Jan;51(1):27-37. doi: 10.1093/jac/dkg031. J Antimicrob Chemother. 2003. PMID: 12493784
-
Transcriptional regulation of the mtrCDE efflux pump operon: importance for Neisseria gonorrhoeae antimicrobial resistance.Microbiology (Reading). 2022 Aug;168(8):001231. doi: 10.1099/mic.0.001231. Microbiology (Reading). 2022. PMID: 35916832 Free PMC article. Review.
-
Genetic organization and regulation of antimicrobial efflux systems possessed by Neisseria gonorrhoeae and Neisseria meningitidis.J Mol Microbiol Biotechnol. 2001 Apr;3(2):219-24. J Mol Microbiol Biotechnol. 2001. PMID: 11321577 Review.
Cited by
-
Molecular mechanisms of antibiotic resistance.Nat Rev Microbiol. 2015 Jan;13(1):42-51. doi: 10.1038/nrmicro3380. Epub 2014 Dec 1. Nat Rev Microbiol. 2015. PMID: 25435309 Review.
-
Characterization of the Neisseria gonorrhoeae Iron and Fur Regulatory Network.J Bacteriol. 2016 Jul 28;198(16):2180-91. doi: 10.1128/JB.00166-16. Print 2016 Aug 15. J Bacteriol. 2016. PMID: 27246574 Free PMC article.
-
Impact of Species Diversity on the Design of RNA-Based Diagnostics for Antibiotic Resistance in Neisseria gonorrhoeae.Antimicrob Agents Chemother. 2019 Jul 25;63(8):e00549-19. doi: 10.1128/AAC.00549-19. Print 2019 Aug. Antimicrob Agents Chemother. 2019. PMID: 31138575 Free PMC article.
-
RND Efflux Pump Induction: A Crucial Network Unveiling Adaptive Antibiotic Resistance Mechanisms of Gram-Negative Bacteria.Antibiotics (Basel). 2024 May 28;13(6):501. doi: 10.3390/antibiotics13060501. Antibiotics (Basel). 2024. PMID: 38927168 Free PMC article. Review.
-
Neisseria gonorrhoeae host adaptation and pathogenesis.Nat Rev Microbiol. 2018 Apr;16(4):226-240. doi: 10.1038/nrmicro.2017.169. Epub 2018 Feb 12. Nat Rev Microbiol. 2018. PMID: 29430011 Free PMC article. Review.
References
-
- Chasteen TG, Fuentes DE, Tantalean JC, Vasquez CC. 2009. Tellurite: history, oxidative stress, and molecular mechanisms of resistance. FEMS Microbiol. Rev. 33:820–832 - PubMed
-
- Cohen MS, et al. 1997. Reduction of concentration of HIV-1 in semen after treatment of urethritis: implications for prevention of sexual transmission of HIV-1. AIDSCAP Malawi Research Group. Lancet 349:1868–1873 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
