

CNS recruitment of CD8+ T lymphocytes specific for a peripheral virus infection triggers neuropathogenesis during polymicrobial challenge
- PMID: 22216008
- PMCID: PMC3245314
- DOI: 10.1371/journal.ppat.1002462
CNS recruitment of CD8+ T lymphocytes specific for a peripheral virus infection triggers neuropathogenesis during polymicrobial challenge
Abstract
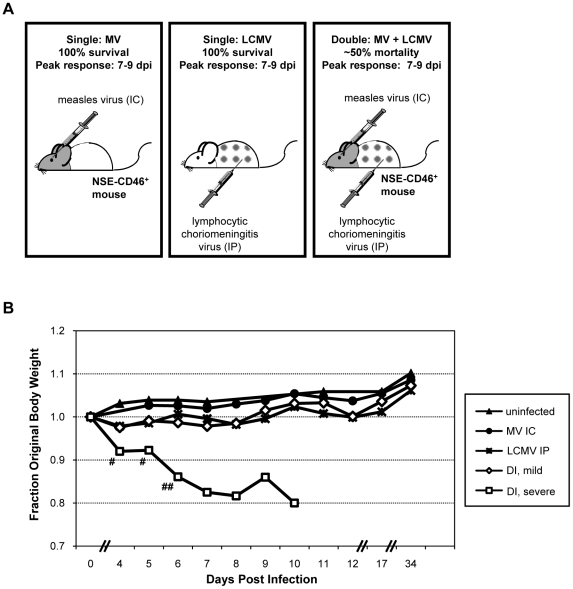
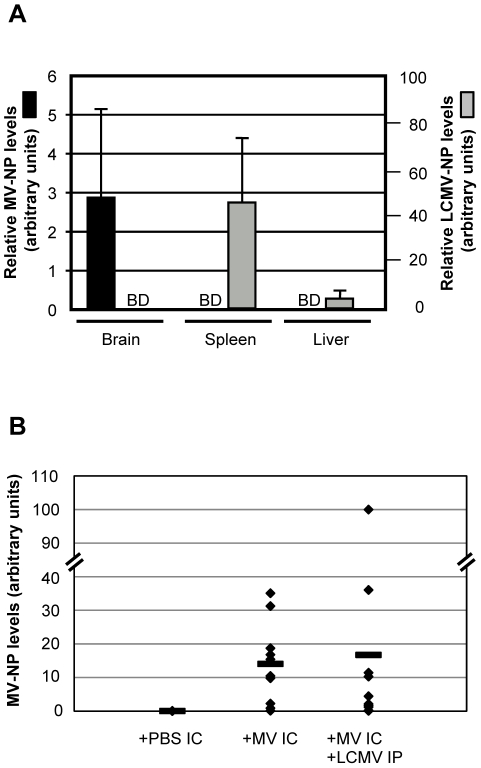
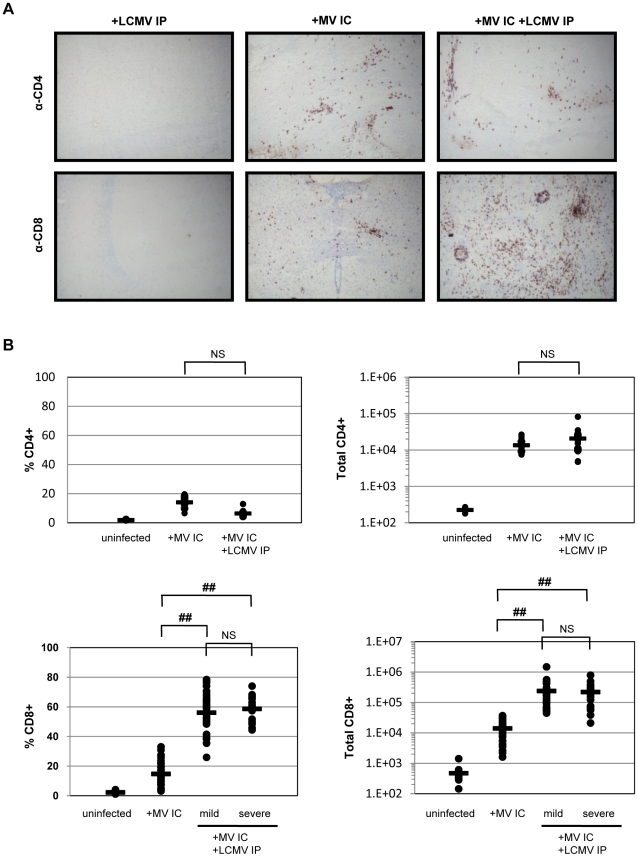
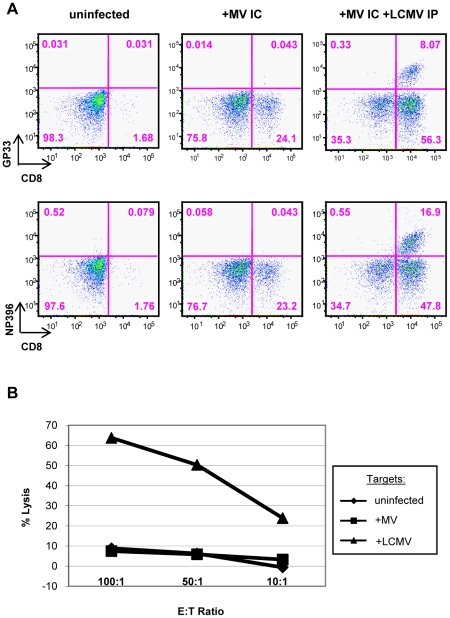
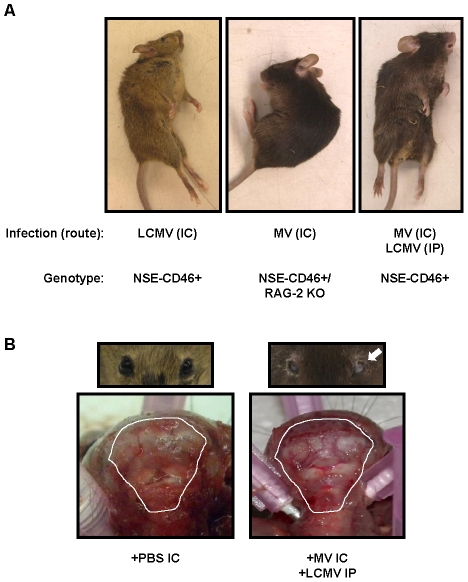
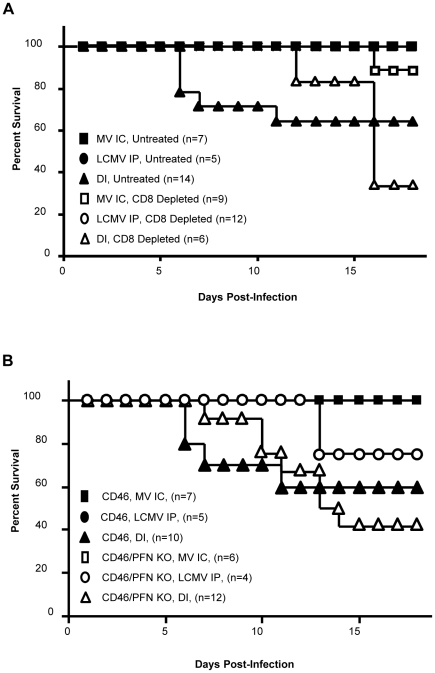
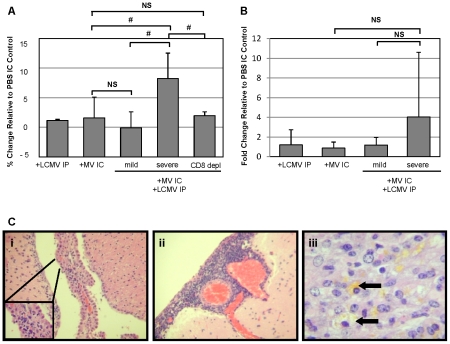
Although viruses have been implicated in central nervous system (CNS) diseases of unknown etiology, including multiple sclerosis and amyotrophic lateral sclerosis, the reproducible identification of viral triggers in such diseases has been largely unsuccessful. Here, we explore the hypothesis that viruses need not replicate in the tissue in which they cause disease; specifically, that a peripheral infection might trigger CNS pathology. To test this idea, we utilized a transgenic mouse model in which we found that immune cells responding to a peripheral infection are recruited to the CNS, where they trigger neurological damage. In this model, mice are infected with both CNS-restricted measles virus (MV) and peripherally restricted lymphocytic choriomeningitis virus (LCMV). While infection with either virus alone resulted in no illness, infection with both viruses caused disease in all mice, with ∼50% dying following seizures. Co-infection resulted in a 12-fold increase in the number of CD8+ T cells in the brain as compared to MV infection alone. Tetramer analysis revealed that a substantial proportion (>35%) of these infiltrating CD8+ lymphocytes were LCMV-specific, despite no detectable LCMV in CNS tissues. Mechanistically, CNS disease was due to edema, induced in a CD8-dependent but perforin-independent manner, and brain herniation, similar to that observed in mice challenged intracerebrally with LCMV. These results indicate that T cell trafficking can be influenced by other ongoing immune challenges, and that CD8+ T cell recruitment to the brain can trigger CNS disease in the apparent absence of cognate antigen. By extrapolation, human CNS diseases of unknown etiology need not be associated with infection with any particular agent; rather, a condition that compromises and activates the blood-brain barrier and adjacent brain parenchyma can render the CNS susceptible to pathogen-independent immune attack.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Efficient T-cell surveillance of the CNS requires expression of the CXC chemokine receptor 3.J Neurosci. 2004 May 19;24(20):4849-58. doi: 10.1523/JNEUROSCI.0123-04.2004. J Neurosci. 2004. PMID: 15152045 Free PMC article.
-
CXCL10 is the key ligand for CXCR3 on CD8+ effector T cells involved in immune surveillance of the lymphocytic choriomeningitis virus-infected central nervous system.J Immunol. 2006 Apr 1;176(7):4235-43. doi: 10.4049/jimmunol.176.7.4235. J Immunol. 2006. PMID: 16547260
-
Protection of CD3 delta knockout mice from lymphocytic choriomeningitis virus-induced immunopathology: implications for viral neuroinvasion.Virology. 2000 Apr 10;269(2):248-56. doi: 10.1006/viro.2000.0224. Virology. 2000. PMID: 10753703
-
Lymphocytic choriomeningitis infection of the central nervous system.Front Biosci. 2008 May 1;13:4529-43. doi: 10.2741/3021. Front Biosci. 2008. PMID: 18508527 Free PMC article. Review.
-
T cells in the central nervous system: the delicate balance between viral clearance and disease.J Infect Dis. 2002 Dec 1;186 Suppl 2(Suppl 2):S145-51. doi: 10.1086/344264. J Infect Dis. 2002. PMID: 12424690 Free PMC article. Review.
Cited by
-
T cell activation triggers reversible inosine-5'-monophosphate dehydrogenase assembly.J Cell Sci. 2018 Sep 5;131(17):jcs223289. doi: 10.1242/jcs.223289. J Cell Sci. 2018. PMID: 30154209 Free PMC article.
-
Viruses and endogenous retroviruses in multiple sclerosis: From correlation to causation.Acta Neurol Scand. 2017 Dec;136(6):606-616. doi: 10.1111/ane.12775. Epub 2017 May 23. Acta Neurol Scand. 2017. PMID: 28542724 Free PMC article. Review.
-
T cells in the brain enhance neonatal mortality during peripheral LCMV infection.PLoS Pathog. 2021 Jan 5;17(1):e1009066. doi: 10.1371/journal.ppat.1009066. eCollection 2021 Jan. PLoS Pathog. 2021. PMID: 33400715 Free PMC article.
-
Adolescent neuroimmune function and its interaction with alcohol.Int Rev Neurobiol. 2022;161:167-208. doi: 10.1016/bs.irn.2021.08.006. Epub 2021 Oct 4. Int Rev Neurobiol. 2022. PMID: 34801169 Free PMC article. Review.
-
Multiple sclerosis: autoimmunity and viruses.Curr Opin Rheumatol. 2013 Jul;25(4):496-501. doi: 10.1097/BOR.0b013e328362004d. Curr Opin Rheumatol. 2013. PMID: 23656710 Free PMC article. Review.
References
-
- Antel JP, Owens T. The attraction of adhesion molecules. Ann Neurol. 1993;34:123–124. - PubMed
-
- Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. - PubMed
-
- Mackay CR. Homing of naive, memory and effector lymphocytes. Curr Opin Immunol. 1993;5:423–427. - PubMed
-
- Chapman TJ, Castrucci MR, Padrick RC, Bradley LM, Topham DJ. Antigen-specific and non-specific CD4+ T cell recruitment and proliferation during influenza infection. Virology. 2005;340:296–306. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
