

A complex genomic rearrangement involving the endothelin 3 locus causes dermal hyperpigmentation in the chicken
- PMID: 22216010
- PMCID: PMC3245302
- DOI: 10.1371/journal.pgen.1002412
A complex genomic rearrangement involving the endothelin 3 locus causes dermal hyperpigmentation in the chicken
Abstract
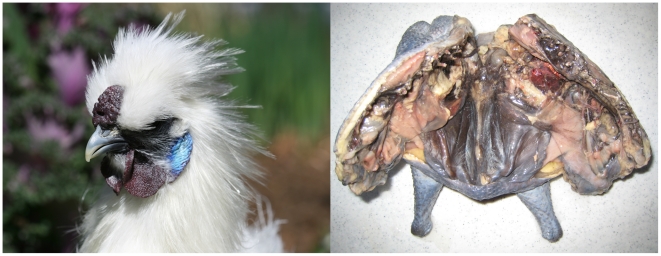
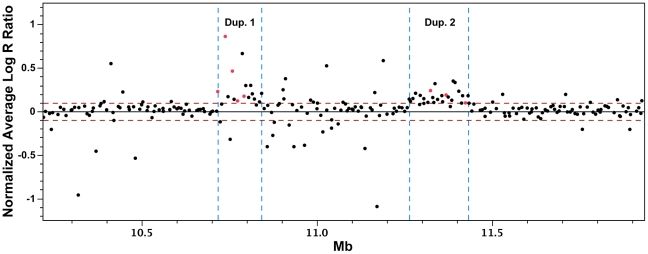
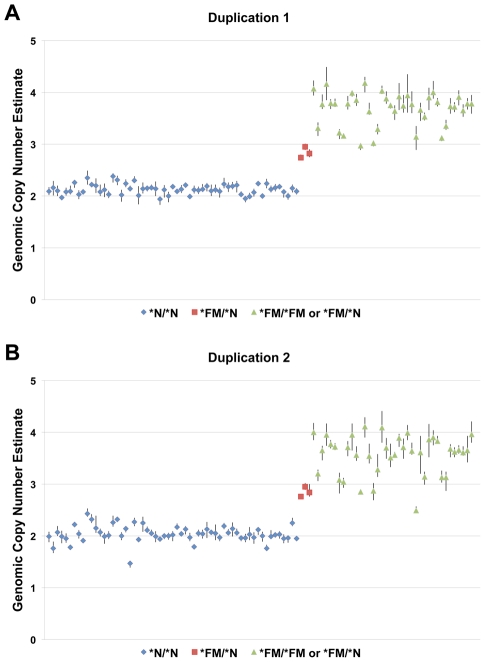
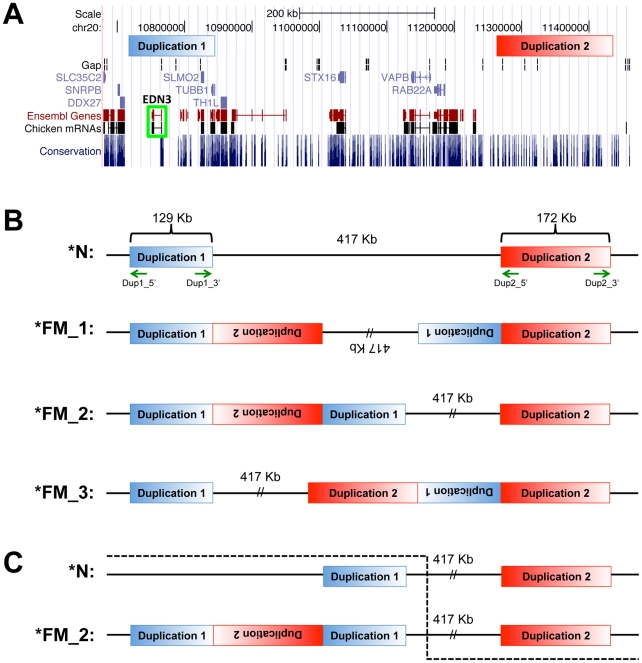
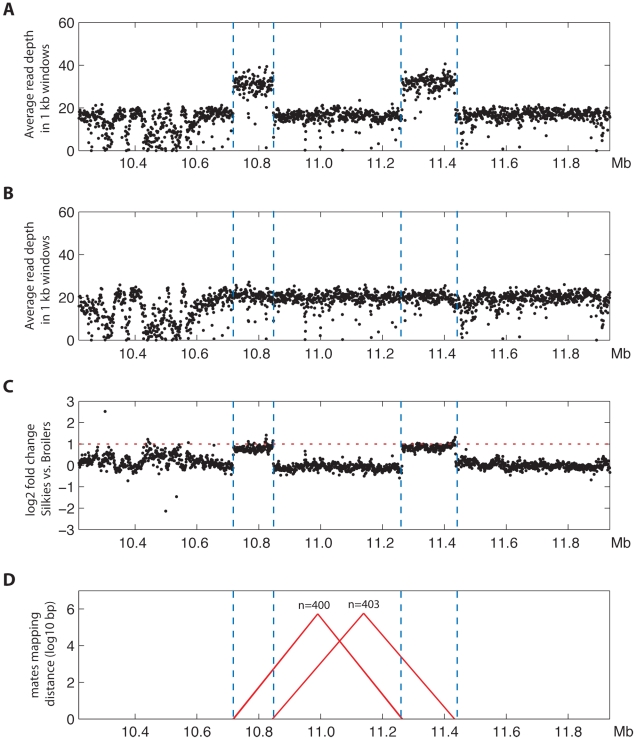
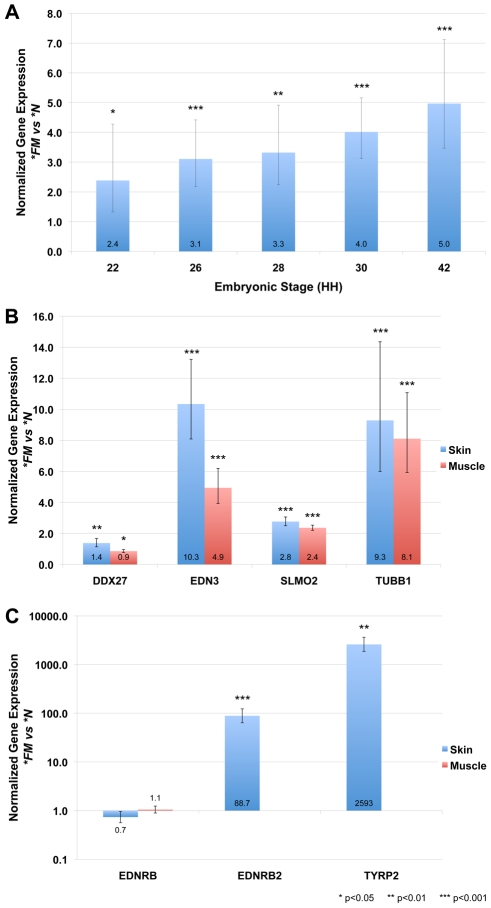
Dermal hyperpigmentation or Fibromelanosis (FM) is one of the few examples of skin pigmentation phenotypes in the chicken, where most other pigmentation variants influence feather color and patterning. The Silkie chicken is the most widespread and well-studied breed displaying this phenotype. The presence of the dominant FM allele results in extensive pigmentation of the dermal layer of skin and the majority of internal connective tissue. Here we identify the causal mutation of FM as an inverted duplication and junction of two genomic regions separated by more than 400 kb in wild-type individuals. One of these duplicated regions contains endothelin 3 (EDN3), a gene with a known role in promoting melanoblast proliferation. We show that EDN3 expression is increased in the developing Silkie embryo during the time in which melanoblasts are migrating, and elevated levels of expression are maintained in the adult skin tissue. We have examined four different chicken breeds from both Asia and Europe displaying dermal hyperpigmentation and conclude that the same structural variant underlies this phenotype in all chicken breeds. This complex genomic rearrangement causing a specific monogenic trait in the chicken illustrates how novel mutations with major phenotypic effects have been reused during breed formation in domestic animals.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Hutt FB. Genetics of the fowl. New York: McGraw-Hill; 1949. pp. 590.
-
- Faraco CD, Vaz SA, Pastor MV, Erickson CA. Hyperpigmentation in the Silkie fowl correlates with abnormal migration of fate-restricted melanoblasts and loss of environmental barrier molecules. Dev Dyn. 2001;220:212–225. - PubMed
-
- Kuklenski J. Uber das Vorkommen und die Verteilung des Pigmentes in den Organen und Geweben bei japanischen seiden Huhnern. Arch Micro Anat Entwickl. 1915;87:1–37.
-
- Smyth JR., Jr . Genetics of plumage, skin and eye pigmentation in chickens; In: Crawford RD, editor. Amsterdam; New York: Elsevier; 1990. pp. 109–167.
-
- Muroya S, Tanabe R, Nakajima I, Chikuni K. Molecular characteristics and site specific distribution of the pigment of the silky fowl. J Vet Med Sci. 2000;62:391–395. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
