

Improved resolution of reef-coral endosymbiont (Symbiodinium) species diversity, ecology, and evolution through psbA non-coding region genotyping
- PMID: 22216157
- PMCID: PMC3247227
- DOI: 10.1371/journal.pone.0029013
Improved resolution of reef-coral endosymbiont (Symbiodinium) species diversity, ecology, and evolution through psbA non-coding region genotyping
Abstract
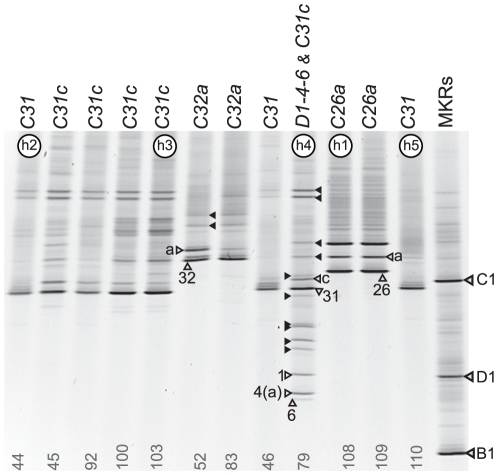
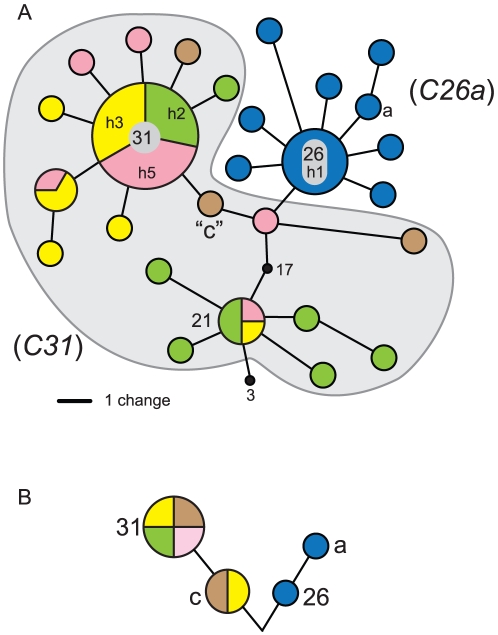
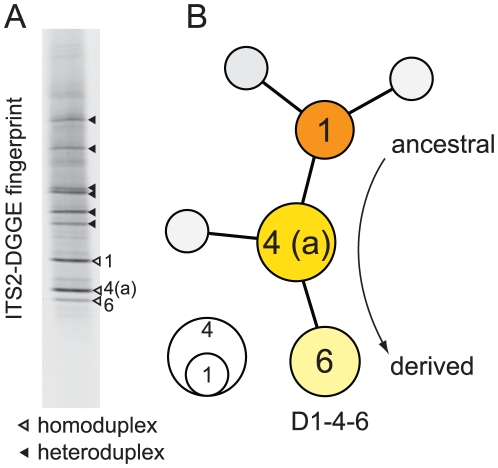
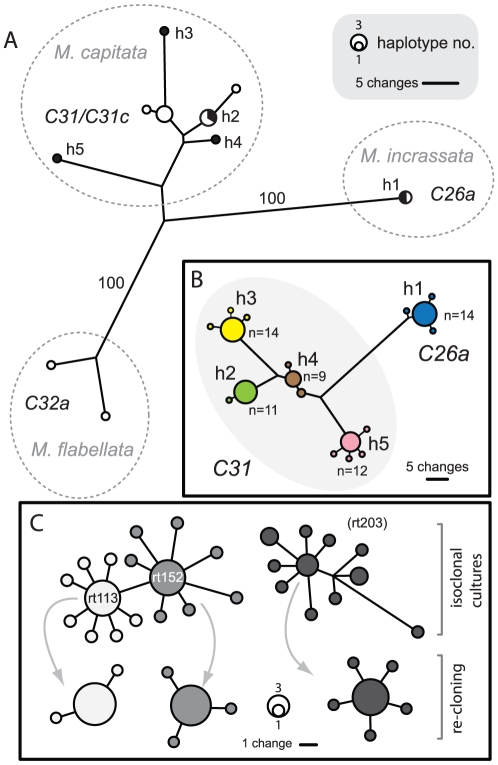
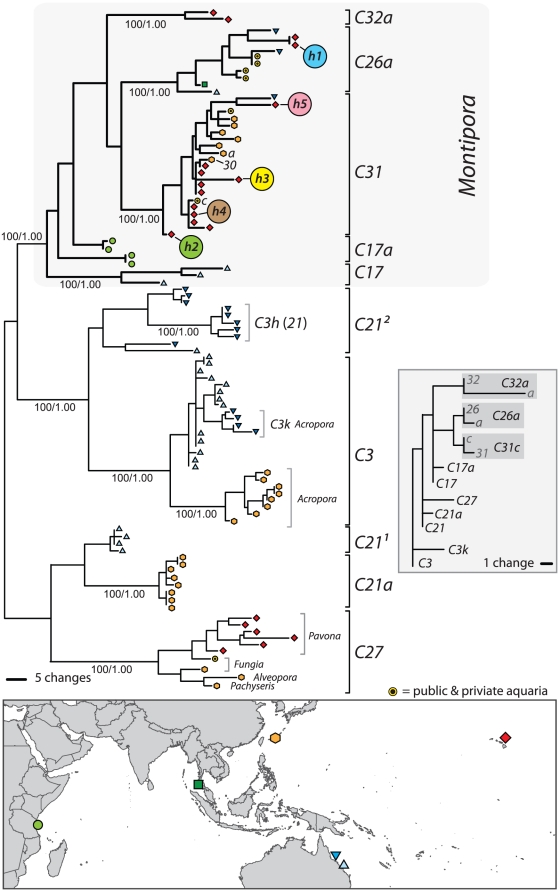
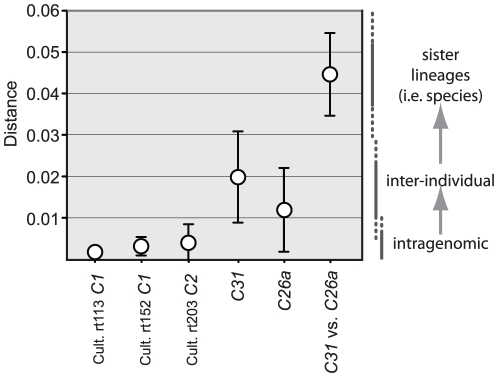
Ribosomal DNA sequence data abounds from numerous studies on the dinoflagellate endosymbionts of corals, and yet the multi-copy nature and intragenomic variability of rRNA genes and spacers confound interpretations of symbiont diversity and ecology. Making consistent sense of extensive sequence variation in a meaningful ecological and evolutionary context would benefit from the application of additional genetic markers. Sequences of the non-coding region of the plastid psbA minicircle (psbA(ncr)) were used to independently examine symbiont genotypic and species diversity found within and between colonies of Hawaiian reef corals in the genus Montipora. A single psbA(ncr) haplotype was recovered in most samples through direct sequencing (~80-90%) and members of the same internal transcribed spacer region 2 (ITS2) type were phylogenetically differentiated from other ITS2 types by substantial psbA(ncr) sequence divergence. The repeated sequencing of bacterially-cloned fragments of psbA(ncr) from samples and clonal cultures often recovered a single numerically common haplotype accompanied by rare, highly-similar, sequence variants. When sequence artifacts of cloning and intragenomic variation are factored out, these data indicate that most colonies harbored one dominant Symbiodinium genotype. The cloning and sequencing of ITS2 DNA amplified from these same samples recovered numerically abundant variants (that are diagnostic of distinct Symbiodinium lineages), but also generated a large amount of sequences comprising PCR/cloning artifacts combined with ancestral and/or rare variants that, if incorporated into phylogenetic reconstructions, confound how small sequence differences are interpreted. Finally, psbA(ncr) sequence data from a broad sampling of Symbiodinium diversity obtained from various corals throughout the Indo-Pacific were concordant with ITS lineage membership (defined by denaturing gradient gel electrophoresis screening), yet exhibited substantially greater sequence divergence and revealed strong phylogeographic structure corresponding to major biogeographic provinces. The detailed genetic resolution provided by psbA(ncr) data brings further clarity to the ecology, evolution, and systematics of symbiotic dinoflagellates.
© 2011 LaJeunesse, Thornhill.
Conflict of interest statement
Figures


References
-
- Rowan R. Diversity and ecology of zooxanthellae on coral reefs. J Phycol. 1998;344:7–1.
-
- LaJeunesse TC. Investigating the biodiversity, ecology, and phylogeny of endosymbiotic dinoflagellates in the genus Symbiodinium using the ITS region: in search of a ‘species’ level marker. J Phycol. 2001;37:866–880.
-
- Baker AC. Flexibility and specificity in coral-algal symbiosis: diversity, ecology, and biogeography of Symbiodinium. Annu Rev Ecol Evol S. 2003;34:661–689.
-
- Coffroth MA, Santos SR. Genetic diversity of symbiotic dinoflagellates in the genus Symbiodinium. Protist. 2005;156:19–34. - PubMed
-
- Rowan R, Powers DA. A molecular genetic classification of zooxanthellae and the evolution of animal-algal symbiosis. Science. 1991;251:1348–1351. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
