

The cytokine release inhibitory drug CRID3 targets ASC oligomerisation in the NLRP3 and AIM2 inflammasomes
- PMID: 22216309
- PMCID: PMC3245271
- DOI: 10.1371/journal.pone.0029539
The cytokine release inhibitory drug CRID3 targets ASC oligomerisation in the NLRP3 and AIM2 inflammasomes
Erratum in
- PLoS One. 2013;8(2). doi:10.1371/annotation/9f221489-155d-4978-a36d-30c51853e438
- PLoS One. 2013;8(7). doi:10.1371/annotation/78d328b9-2c8c-4978-84b1-7e6a0b12ada1. Robertson, Avril [added]; Butler, Mark [added]; Cooper, Matthew [added]
Abstract
Background: The Inflammasomes are multi-protein complexes that regulate caspase-1 activation and the production of the pro-inflammatory cytokine IL-1β. Previous studies identified a class of diarylsulfonylurea containing compounds called Cytokine Release Inhibitory Drugs (CRIDs) that inhibited the post-translational processing of IL-1β. Further work identified Glutathione S-Transferase Omega 1 (GSTO1) as a possible target of these CRIDs. This study aimed to investigate the mechanism of the inhibitory activity of the CRID CP-456,773 (termed CRID3) in light of recent advances in the area of inflammasome activation, and to clarify the potential role of GSTO1 in the regulation of IL-1β production.
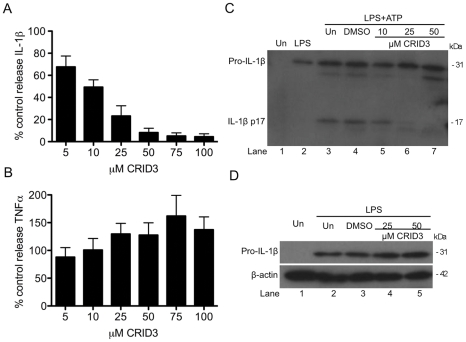
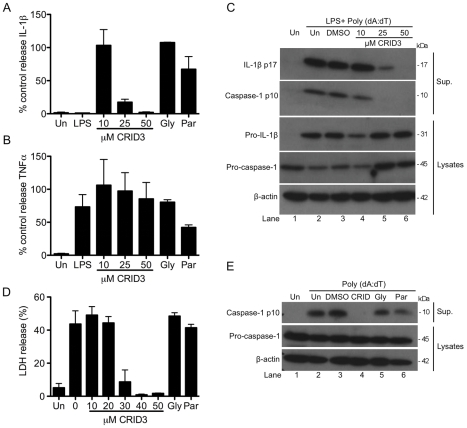
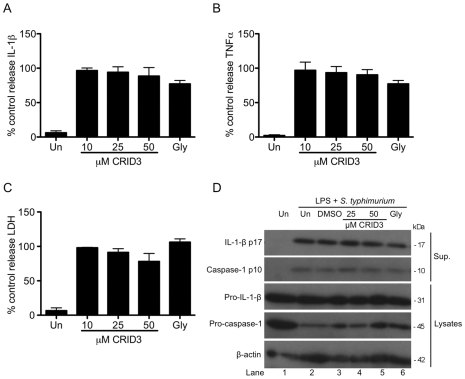
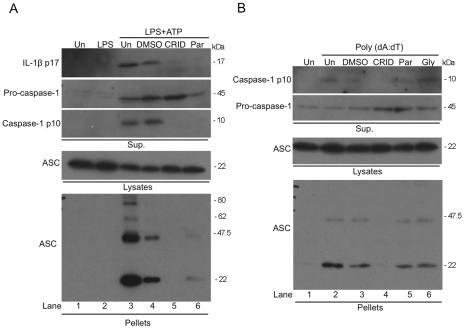
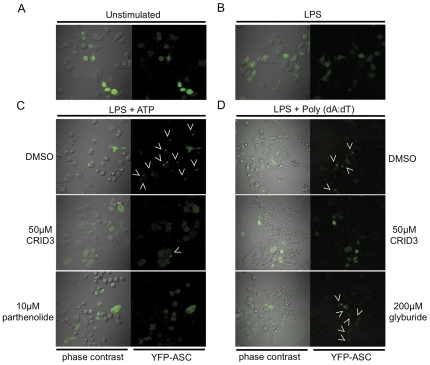
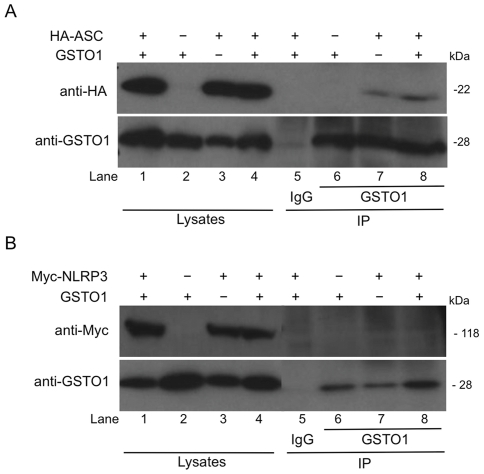
Methodology and results: In murine bone marrow derived macrophages, CRID3 inhibited IL-1β secretion and caspase 1 processing in response to stimulation of NLRP3 and AIM2 but not NLRC4. CRID3 also prevented AIM2 dependent pyroptosis in contrast to the NLRP3 inhibitors glyburide and parthenolide, which do not inhibit AIM2 activation. Confocal microscopy and Western blotting assays indicated that CRID3 inhibited the formation of ASC complexes or 'specks' in response to NLRP3 and AIM2 stimulation. Co-immunoprecipitation assays show that GSTO1 interacted with ASC.
Significance: These results identify CRID3 as a novel inhibitor of the NLRP3 and AIM2 inflammasomes and provide an insight into the mechanism of action of this small molecule. In addition GSTO1 may be a component of the inflammasome that is required for ASC complex formation.
© 2011 Coll, O’Neill.
Conflict of interest statement
Figures
References
-
- Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140:821–832. - PubMed
-
- Keller M, Ruegg A, Werner S, Beer HD. Active caspase-1 is a regulator of unconventional protein secretion. Cell. 2008;132:818–831. - PubMed
-
- Agostini L, Martinon F, Burns K, McDermott MF, Hawkins PN, et al. NALP3 forms an IL-1beta-processing inflammasome with increased activity in Muckle-Wells autoinflammatory disorder. Immunity. 2004;20:319–325. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
