

Endogenous Rho-kinase signaling maintains synaptic strength by stabilizing the size of the readily releasable pool of synaptic vesicles
- PMID: 22219271
- PMCID: PMC6621323
- DOI: 10.1523/JNEUROSCI.3215-11.2012
Endogenous Rho-kinase signaling maintains synaptic strength by stabilizing the size of the readily releasable pool of synaptic vesicles
Abstract
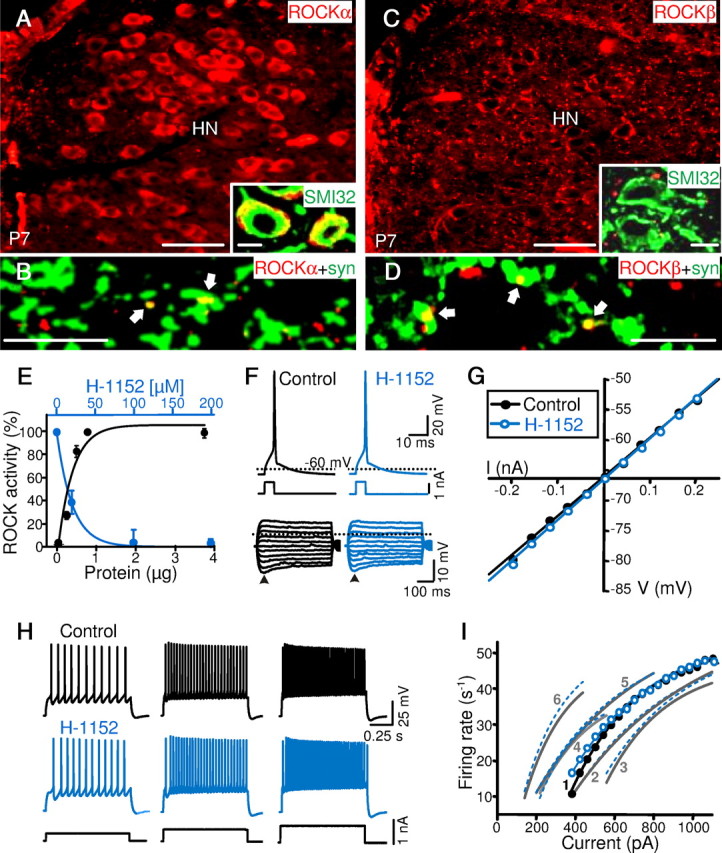
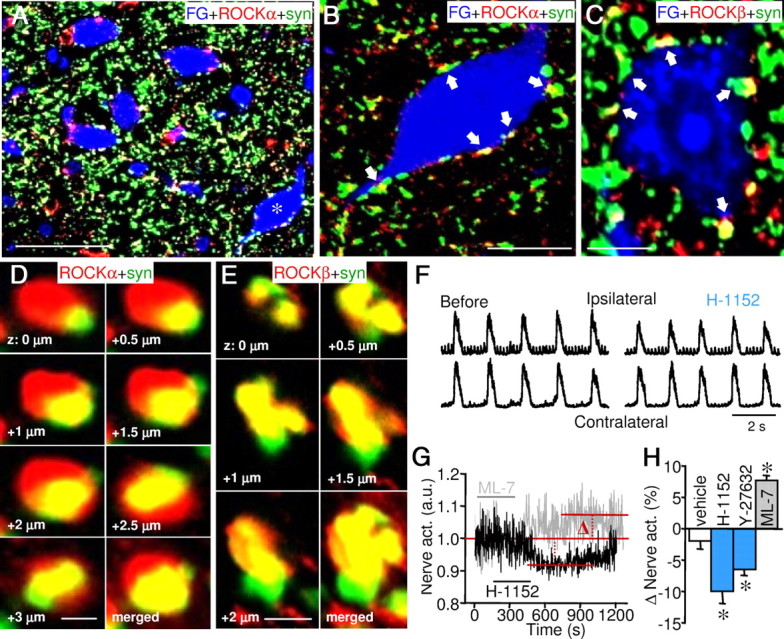
Rho-associated kinase (ROCK) regulates neural cell migration, proliferation and survival, dendritic spine morphology, and axon guidance and regeneration. There is, however, little information about whether ROCK modulates the electrical activity and information processing of neuronal circuits. At neonatal stage, ROCKα is expressed in hypoglossal motoneurons (HMNs) and in their afferent inputs, whereas ROCKβ is found in synaptic terminals on HMNs, but not in their somata. Inhibition of endogenous ROCK activity in neonatal rat brainstem slices failed to modulate intrinsic excitability of HMNs, but strongly attenuated the strength of their glutamatergic and GABAergic synaptic inputs. The mechanism acts presynaptically to reduce evoked neurotransmitter release. ROCK inhibition increased myosin light chain (MLC) phosphorylation, which is known to trigger actomyosin contraction, and reduced the number of synaptic vesicles docked to active zones in excitatory boutons. Functional and ultrastructural changes induced by ROCK inhibition were fully prevented/reverted by MLC kinase (MLCK) inhibition. Furthermore, ROCK inhibition drastically reduced the phosphorylated form of p21-associated kinase (PAK), which directly inhibits MLCK. We conclude that endogenous ROCK activity is necessary for the normal performance of motor output commands, because it maintains afferent synaptic strength, by stabilizing the size of the readily releasable pool of synaptic vesicles. The mechanism of action involves a tonic inhibition of MLCK, presumably through PAK phosphorylation. This mechanism might be present in adults since unilateral microinjection of ROCK or MLCK inhibitors into the hypoglossal nucleus reduced or increased, respectively, whole XIIth nerve activity.
Figures







References
-
- Anliker B, Chun J. Cell surface receptors in lysophospholipid signaling. Semin Cell Dev Biol. 2004;15:457–465. - PubMed
-
- Bähler M, Greengard P. Synapsin I bundles F-actin in a phosphorylation-dependent manner. Nature. 1987;326:704–707. - PubMed
-
- Benarroch EE. Rho GTPases: role in dendrite and axonal growth, mental retardation, and axonal regeneration. Neurology. 2007;68:1315–1318. - PubMed
-
- Burridge K, Wennerberg K. Rho and Rac take center stage. Cell. 2004;116:167–179. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources