

Transgenic mouse model expressing the caspase 6 fragment of mutant huntingtin
- PMID: 22219281
- PMCID: PMC3306223
- DOI: 10.1523/JNEUROSCI.1305-11.2012
Transgenic mouse model expressing the caspase 6 fragment of mutant huntingtin
Abstract
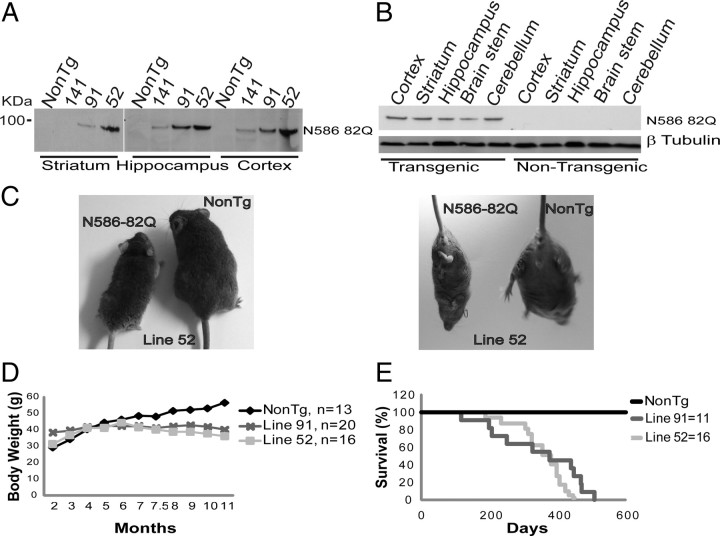
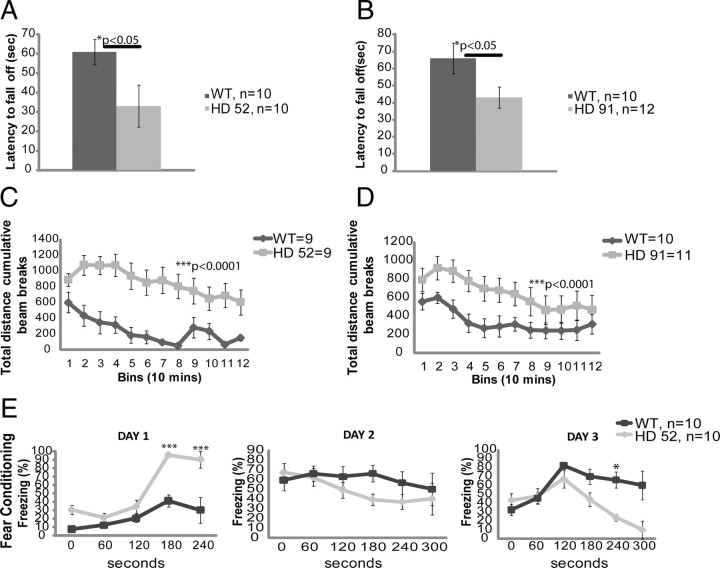
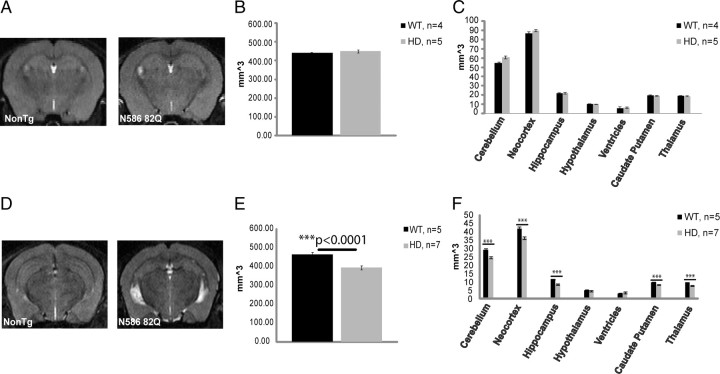
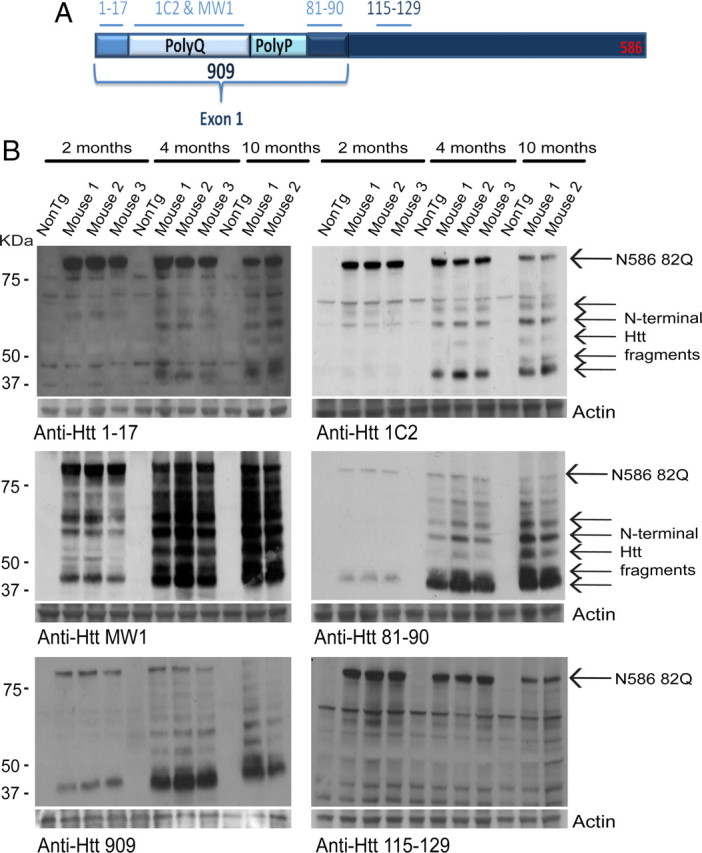
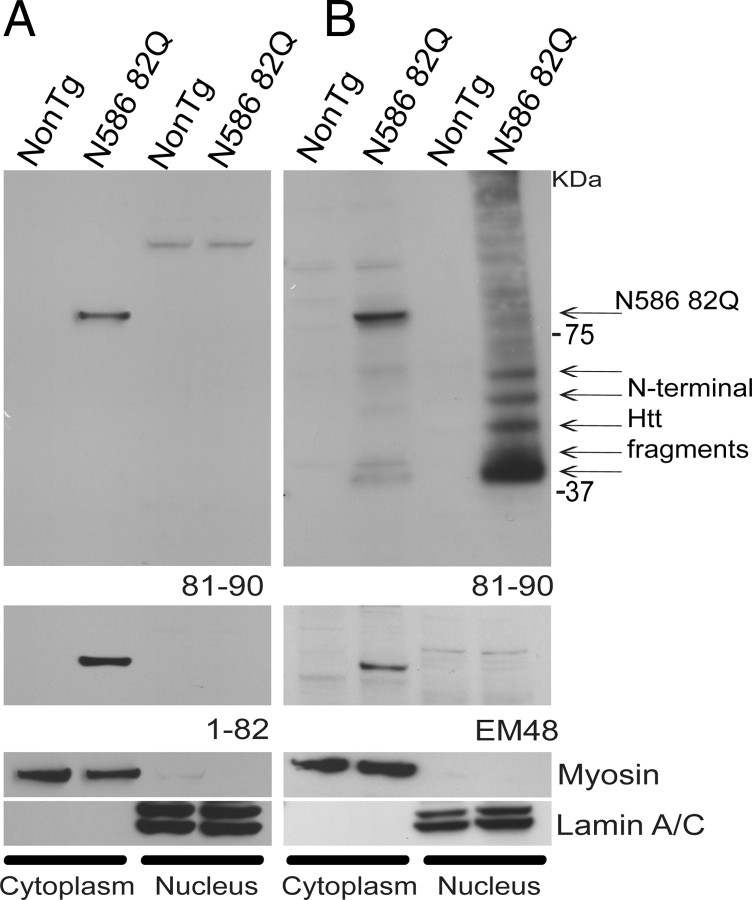
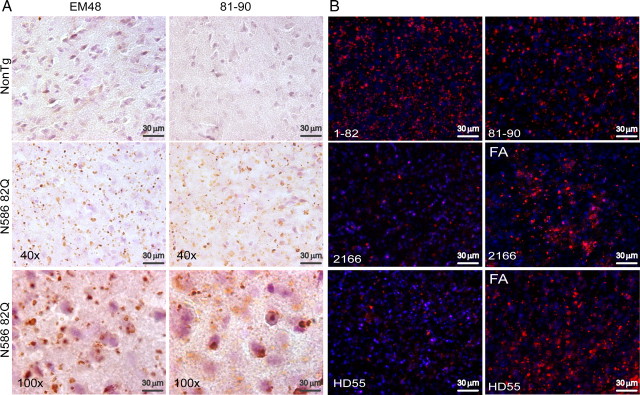
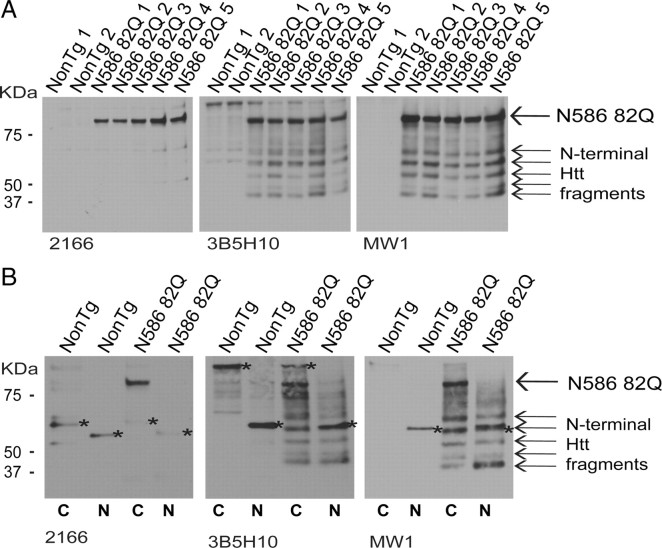
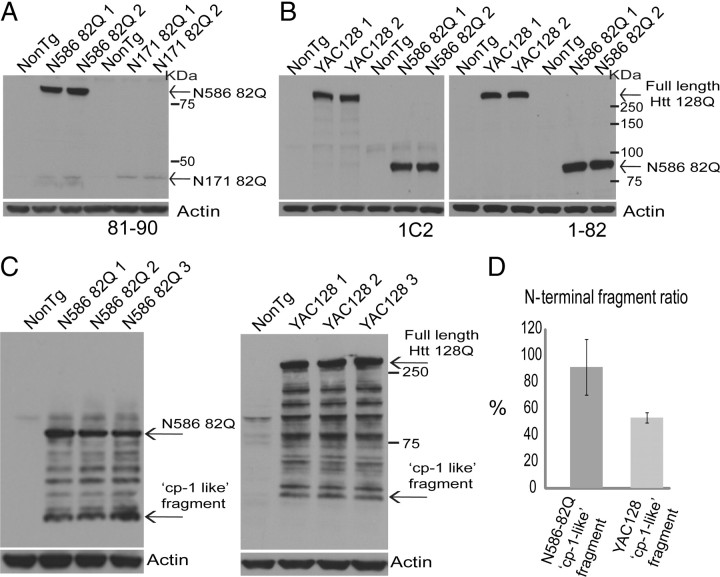
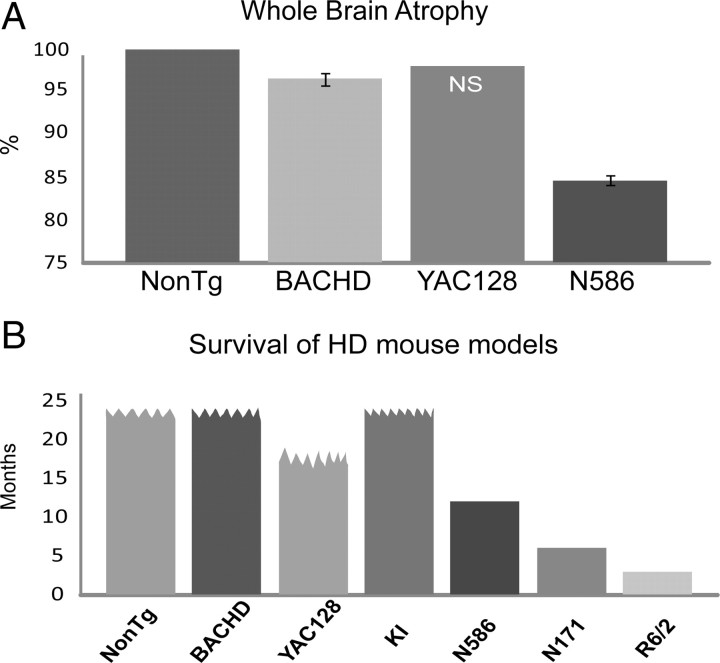
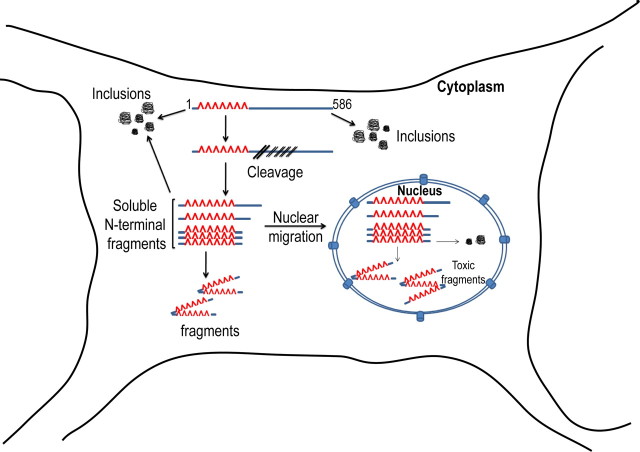
Huntington's disease (HD) is caused by a polyglutamine expansion in the Huntingtin (Htt) protein. Proteolytic cleavage of Htt into toxic N-terminal fragments is believed to be a key aspect of pathogenesis. The best characterized putative cleavage event is at amino acid 586, hypothesized to be mediated by caspase 6. A corollary of the caspase 6 cleavage hypothesis is that the caspase 6 fragment should be a toxic fragment. To test this hypothesis, and further characterize the role of this fragment, we have generated transgenic mice expressing the N-terminal 586 aa of Htt with a polyglutamine repeat length of 82 (N586-82Q), under the control of the prion promoter. N586-82Q mice show a clear progressive rotarod deficit by 4 months of age, and are hyperactive starting at 5 months, later changing to hypoactivity before early mortality. MRI studies reveal widespread brain atrophy, and histologic studies demonstrate an abundance of Htt aggregates, mostly cytoplasmic, which are predominantly composed of the N586-82Q polypeptide. Smaller soluble N-terminal fragments appear to accumulate over time, peaking at 4 months, and are predominantly found in the nuclear fraction. This model appears to have a phenotype more severe than current full-length Htt models, but less severe than HD mouse models expressing shorter Htt fragments. These studies suggest that the caspase 6 fragment may be a transient intermediate, that fragment size is a factor contributing to the rate of disease progression, and that short soluble nuclear fragments may be most relevant to pathogenesis.
Figures
References
-
- Cornett J, Cao F, Wang CE, Ross CA, Bates GP, Li SH, Li XJ. Polyglutamine expansion of huntingtin impairs its nuclear export. Nat Genet. 2005;37:198–204. - PubMed
-
- DiFiglia M, Sapp E, Chase KO, Davies SW, Bates GP, Vonsattel JP, Aronin N. Aggregation of huntingtin in neuronal intranuclear inclusions and dystrophic neurites in brain. Science. 1997;277:1990–1993. - PubMed
-
- Graham RK, Deng Y, Slow EJ, Haigh B, Bissada N, Lu G, Pearson J, Shehadeh J, Bertram L, Murphy Z, Warby SC, Doty CN, Roy S, Wellington CL, Leavitt BR, Raymond LA, Nicholson DW, Hayden MR. Cleavage at the caspase-6 site is required for neuronal dysfunction and degeneration due to mutant huntingtin. Cell. 2006;125:1179–1191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases