

Adenosine enhances sweet taste through A2B receptors in the taste bud
- PMID: 22219293
- PMCID: PMC3566648
- DOI: 10.1523/JNEUROSCI.4070-11.2012
Adenosine enhances sweet taste through A2B receptors in the taste bud
Abstract
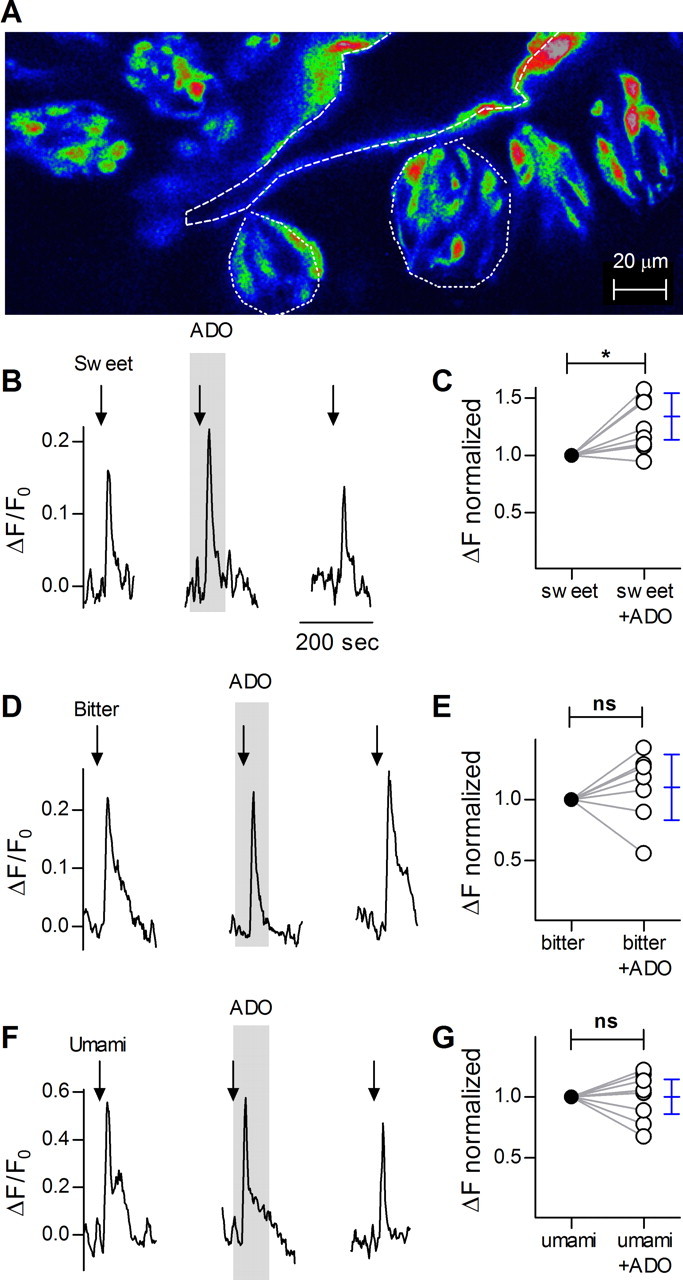
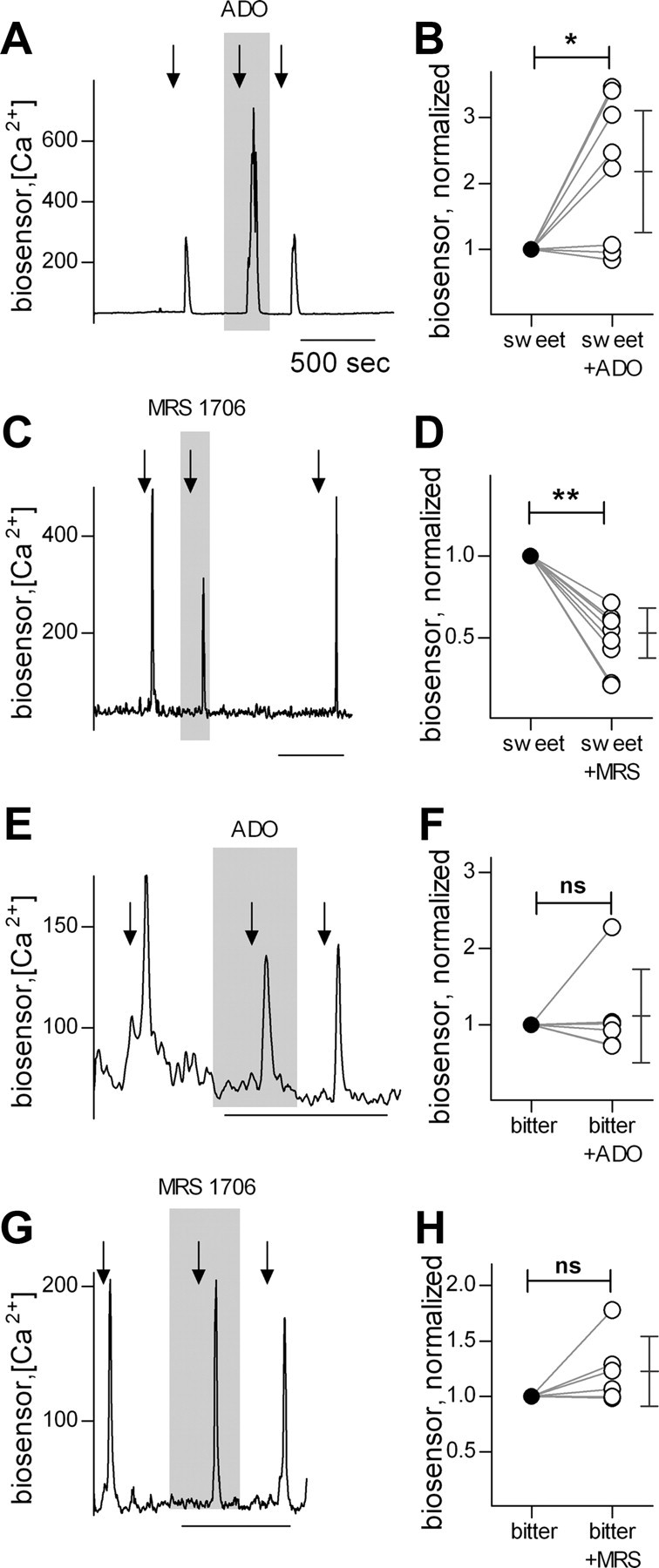
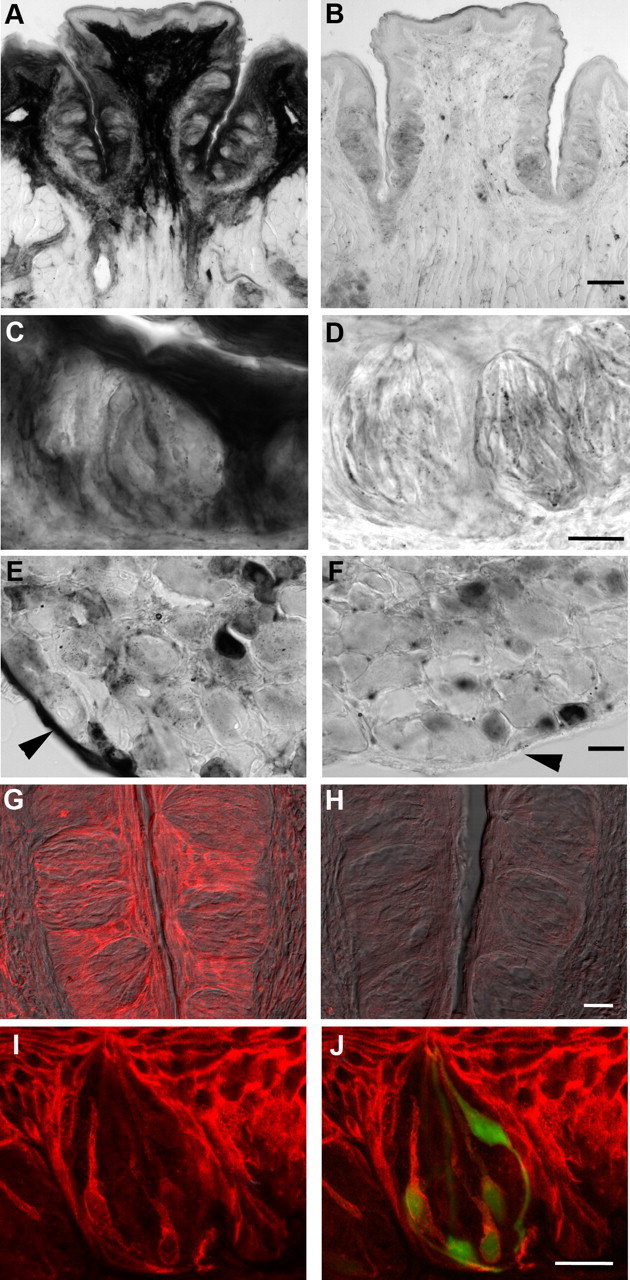
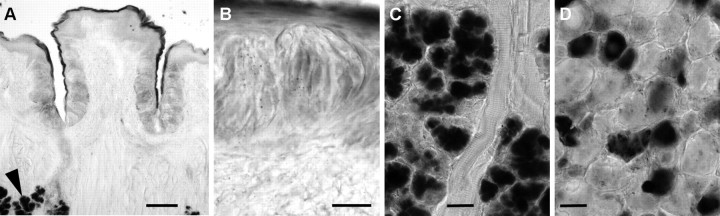
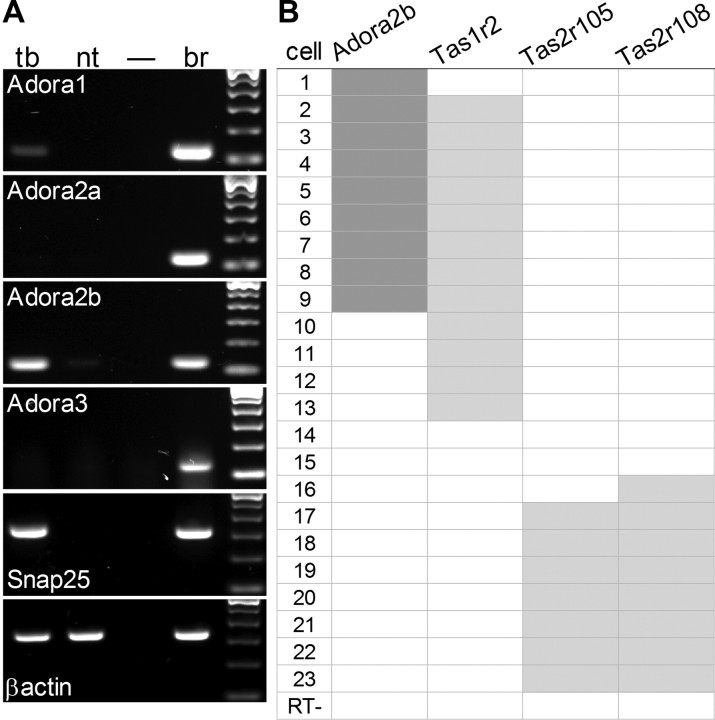
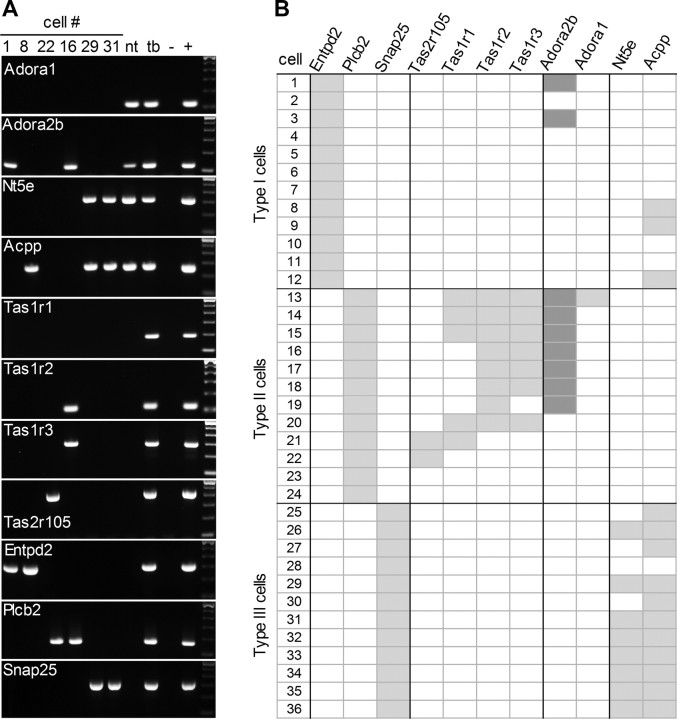
Mammalian taste buds use ATP as a neurotransmitter. Taste Receptor (type II) cells secrete ATP via gap junction hemichannels into the narrow extracellular spaces within a taste bud. This ATP excites primary sensory afferent fibers and also stimulates neighboring taste bud cells. Here we show that extracellular ATP is enzymatically degraded to adenosine within mouse vallate taste buds and that this nucleoside acts as an autocrine neuromodulator to selectively enhance sweet taste. In Receptor cells in a lingual slice preparation, Ca(2+) mobilization evoked by focally applied artificial sweeteners was significantly enhanced by adenosine (50 μM). Adenosine had no effect on bitter or umami taste responses, and the nucleoside did not affect Presynaptic (type III) taste cells. We also used biosensor cells to measure transmitter release from isolated taste buds. Adenosine (5 μM) enhanced ATP release evoked by sweet but not bitter taste stimuli. Using single-cell reverse transcriptase (RT)-PCR on isolated vallate taste cells, we show that many Receptor cells express the adenosine receptor, Adora2b, while Presynaptic (type III) and Glial-like (type I) cells seldom do. Furthermore, Adora2b receptors are significantly associated with expression of the sweet taste receptor subunit, Tas1r2. Adenosine is generated during taste stimulation mainly by the action of the ecto-5'-nucleotidase, NT5E, and to a lesser extent, prostatic acid phosphatase. Both these ecto-nucleotidases are expressed by Presynaptic cells, as shown by single-cell RT-PCR, enzyme histochemistry, and immunofluorescence. Our findings suggest that ATP released during taste reception is degraded to adenosine to exert positive modulation particularly on sweet taste.
Figures
Similar articles
-
A2BR adenosine receptor modulates sweet taste in circumvallate taste buds.PLoS One. 2012;7(1):e30032. doi: 10.1371/journal.pone.0030032. Epub 2012 Jan 10. PLoS One. 2012. PMID: 22253866 Free PMC article.
-
Calcitonin Gene-Related Peptide Reduces Taste-Evoked ATP Secretion from Mouse Taste Buds.J Neurosci. 2015 Sep 16;35(37):12714-24. doi: 10.1523/JNEUROSCI.0100-15.2015. J Neurosci. 2015. PMID: 26377461 Free PMC article.
-
Leptin's effect on taste bud calcium responses and transmitter secretion.Chem Senses. 2015 May;40(4):217-22. doi: 10.1093/chemse/bju066. Epub 2014 Dec 23. Chem Senses. 2015. PMID: 25537017 Free PMC article.
-
Taste buds as peripheral chemosensory processors.Semin Cell Dev Biol. 2013 Jan;24(1):71-9. doi: 10.1016/j.semcdb.2012.12.002. Epub 2012 Dec 20. Semin Cell Dev Biol. 2013. PMID: 23261954 Free PMC article. Review.
-
The Role of ATP and Purinergic Receptors in Taste Signaling.Handb Exp Pharmacol. 2022;275:91-107. doi: 10.1007/164_2021_518. Handb Exp Pharmacol. 2022. PMID: 34435233 Free PMC article. Review.
Cited by
-
Synaptic communication and signal processing among sensory cells in taste buds.J Physiol. 2014 Aug 15;592(16):3387-92. doi: 10.1113/jphysiol.2013.269837. Epub 2014 Mar 24. J Physiol. 2014. PMID: 24665098 Free PMC article.
-
A permeability barrier surrounds taste buds in lingual epithelia.Am J Physiol Cell Physiol. 2015 Jan 1;308(1):C21-32. doi: 10.1152/ajpcell.00157.2014. Epub 2014 Sep 10. Am J Physiol Cell Physiol. 2015. PMID: 25209263 Free PMC article.
-
A2BR adenosine receptor modulates sweet taste in circumvallate taste buds.PLoS One. 2012;7(1):e30032. doi: 10.1371/journal.pone.0030032. Epub 2012 Jan 10. PLoS One. 2012. PMID: 22253866 Free PMC article.
-
Re-rolling treatment in the fermentation process improves the taste and liquor color qualities of black tea.Food Chem X. 2024 Jan 20;21:101143. doi: 10.1016/j.fochx.2024.101143. eCollection 2024 Mar 30. Food Chem X. 2024. PMID: 38312489 Free PMC article.
-
Taste Bud Connectome: Implications for Taste Information Processing.J Neurosci. 2022 Feb 2;42(5):804-816. doi: 10.1523/JNEUROSCI.0838-21.2021. Epub 2021 Dec 7. J Neurosci. 2022. PMID: 34876471 Free PMC article.
References
-
- Baldwin SA, Beal PR, Yao SY, King AE, Cass CE, Young JD. The equilibrative nucleoside transporter family, SLC29. Pflugers Arch. 2004;447:735–743. - PubMed
-
- Chandrashekar J, Mueller KL, Hoon MA, Adler E, Feng L, Guo W, Zuker CS, Ryba NJ. T2Rs function as bitter taste receptors. Cell. 2000;100:703–711. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous