

Single-molecule imaging of translational output from individual RNA granules in neurons
- PMID: 22219377
- PMCID: PMC3290649
- DOI: 10.1091/mbc.E11-07-0622
Single-molecule imaging of translational output from individual RNA granules in neurons
Abstract
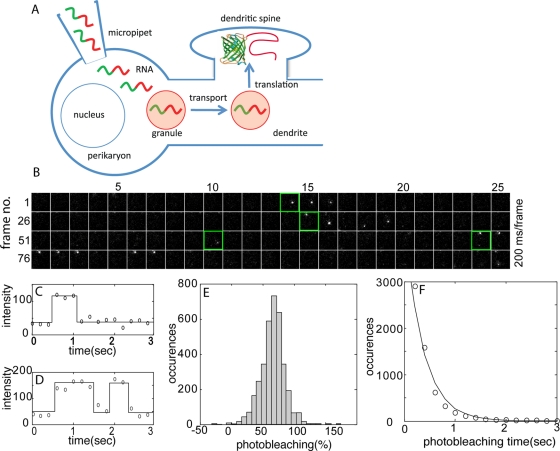
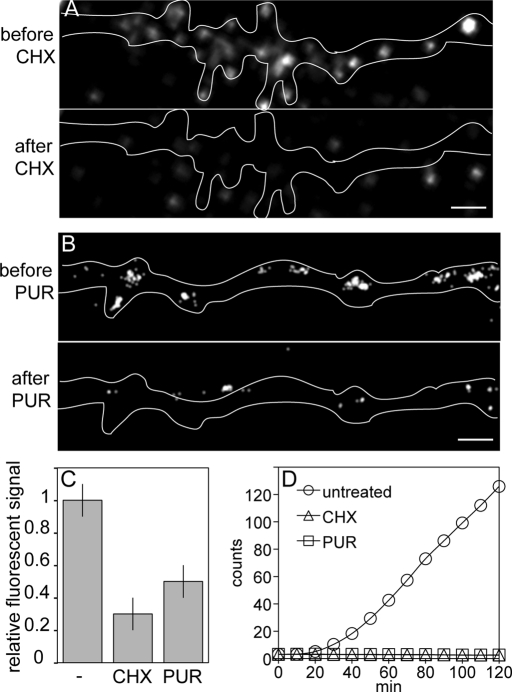
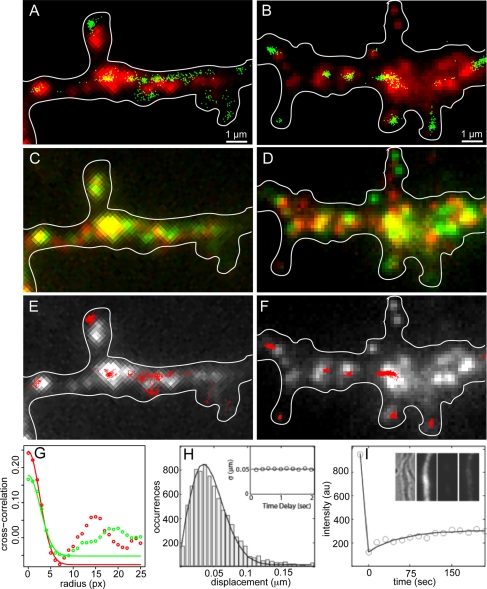
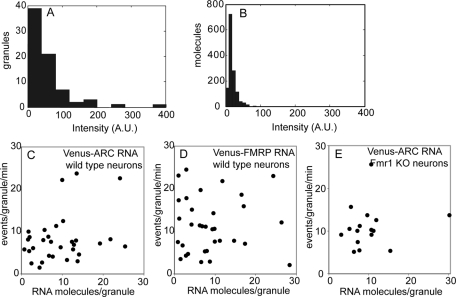
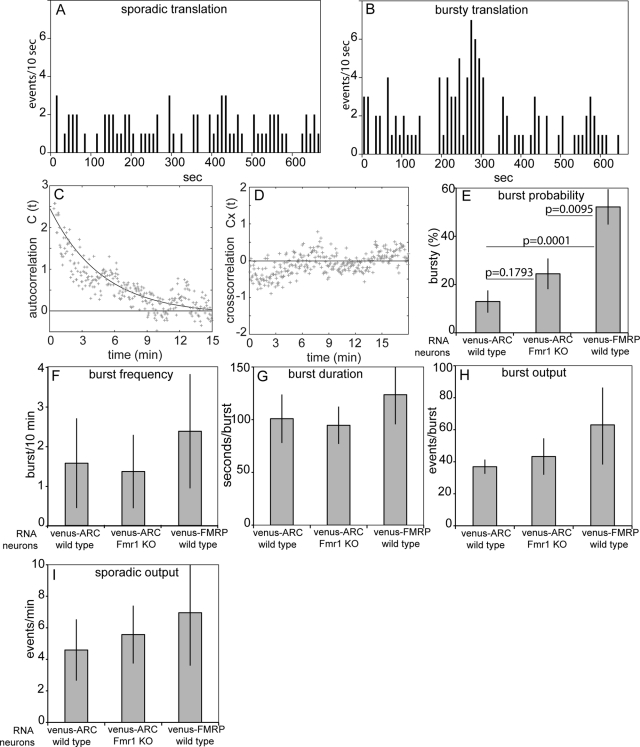
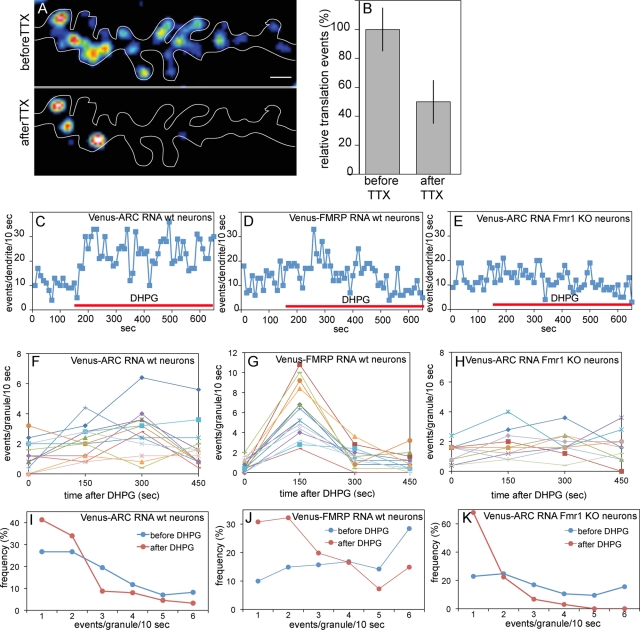
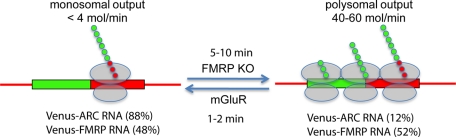
Dendritic RNAs are localized and translated in RNA granules. Here we use single-molecule imaging to count the number of RNA molecules in each granule and to record translation output from each granule using Venus fluorescent protein as a reporter. For RNAs encoding activity-regulated cytoskeletal-associated protein (ARC) or fragile X mental retardation protein (FMRP), translation events are spatially clustered near individual granules, and translational output from individual granules is either sporadic or bursty. The probability of bursty translation is greater for Venus-FMRP RNA than for Venus-ARC RNA and is increased in Fmr1-knockout neurons compared to wild-type neurons. Dihydroxyphenylglycine (DHPG) increases the rate of sporadic translation and decreases bursty translation for Venus-FMRP and Venus-ARC RNAs. Single-molecule imaging of translation in individual granules provides new insight into molecular, spatial, and temporal regulation of translation in granules.
Figures
Similar articles
-
CGG Repeats in the 5'UTR of FMR1 RNA Regulate Translation of Other RNAs Localized in the Same RNA Granules.PLoS One. 2016 Dec 22;11(12):e0168204. doi: 10.1371/journal.pone.0168204. eCollection 2016. PLoS One. 2016. PMID: 28005950 Free PMC article.
-
Evidence for a fragile X mental retardation protein-mediated translational switch in metabotropic glutamate receptor-triggered Arc translation and long-term depression.J Neurosci. 2012 Apr 25;32(17):5924-36. doi: 10.1523/JNEUROSCI.4650-11.2012. J Neurosci. 2012. PMID: 22539853 Free PMC article.
-
Transport of fragile X mental retardation protein via granules in neurites of PC12 cells.Mol Cell Biol. 2002 Dec;22(23):8332-41. doi: 10.1128/MCB.22.23.8332-8341.2002. Mol Cell Biol. 2002. PMID: 12417734 Free PMC article.
-
Cataloguing and Selection of mRNAs Localized to Dendrites in Neurons and Regulated by RNA-Binding Proteins in RNA Granules.Biomolecules. 2020 Jan 22;10(2):167. doi: 10.3390/biom10020167. Biomolecules. 2020. PMID: 31978946 Free PMC article. Review.
-
mRNPs, polysomes or granules: FMRP in neuronal protein synthesis.Curr Opin Neurobiol. 2006 Jun;16(3):265-9. doi: 10.1016/j.conb.2006.05.010. Epub 2006 May 16. Curr Opin Neurobiol. 2006. PMID: 16707258 Review.
Cited by
-
Molecular physiology of Arc/Arg3.1: The oligomeric state hypothesis of synaptic plasticity.Acta Physiol (Oxf). 2022 Nov;236(3):e13886. doi: 10.1111/apha.13886. Epub 2022 Sep 20. Acta Physiol (Oxf). 2022. PMID: 36073248 Free PMC article. Review.
-
Quantifying Single mRNA Translation Kinetics in Living Cells.Cold Spring Harb Perspect Biol. 2018 Nov 1;10(11):a032078. doi: 10.1101/cshperspect.a032078. Cold Spring Harb Perspect Biol. 2018. PMID: 30385605 Free PMC article. Review.
-
Single Molecule Translation Imaging Visualizes the Dynamics of Local β-Actin Synthesis in Retinal Axons.Sci Rep. 2017 Apr 6;7(1):709. doi: 10.1038/s41598-017-00695-7. Sci Rep. 2017. PMID: 28386060 Free PMC article.
-
mRNA Targeting, Transport and Local Translation in Eukaryotic Cells: From the Classical View to a Diversity of New Concepts.Mol Biol. 2021;55(4):507-537. doi: 10.1134/S0026893321030080. Epub 2021 May 30. Mol Biol. 2021. PMID: 34092811 Free PMC article.
-
Dendritic protein synthesis in the normal and diseased brain.Neuroscience. 2013 Mar 1;232:106-27. doi: 10.1016/j.neuroscience.2012.12.003. Epub 2012 Dec 20. Neuroscience. 2013. PMID: 23262237 Free PMC article. Review.
References
-
- Antar LN, Dictenberg JB, Plociniak M, Afroz R, Bassell GJ. Localization of FMRP-associated mRNA granules and requirement of microtubules for activity-dependent trafficking in hippocampal neurons. Genes Brain Behav. 2005;4:350–359. - PubMed
-
- Bourne JN, Sorra KE, Hurlburt J, Harris KM. Polyribosomes are increased in spines of CA1 dendrites 2 h after the induction of LTP in mature rat hippocampal slices. Hippocampus. 2007;17:1–4. - PubMed
-
- Castillo PE, Francesconi A, Carroll RC. The ups and downs of translation-dependent plasticity. Neuron. 2008;59:1–3. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
