

Review
doi: 10.1002/9781118105771.ch9.
Transglutaminase-mediated remodeling of the human erythrocyte membrane skeleton: relevance for erythrocyte diseases with shortened cell lifespan
Affiliations
- PMID: 22220479
- PMCID: PMC3313834
- DOI: 10.1002/9781118105771.ch9
Item in Clipboard
Review
Transglutaminase-mediated remodeling of the human erythrocyte membrane skeleton: relevance for erythrocyte diseases with shortened cell lifespan
Adv Enzymol Relat Areas Mol Biol.
2011.
No abstract available
Figures
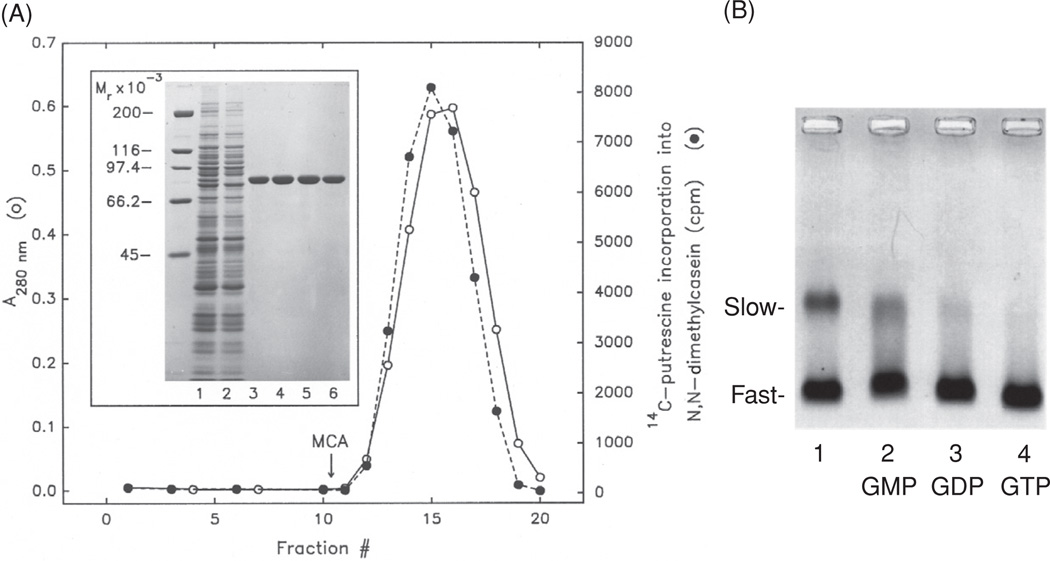
(A) Affinity purification of TG2 by single passage of the hemoglobin-depleted lysate of hRBCs through a column of the 42-kDa gelatin-binding fragment of human fibronectin. The Hb-depleted cell lysate was applied to the affinity column. After extensive washing, the tightly held enzyme was eluted with 0.25% monochloroacetic acid (MCA). Fractions were neutralized and analyzed for protein (left ordinate, open circle) and enzyme activity (right ordinate, closed circle). Inset: eluted samples were examined by SDS-PAGE, stained with Coomassie blue R. Lanes: 1, hemoglobin-depleted lysate; 2, nonretained material passing through the column; 3–6, fractions 14–17 eluted with MCA. Molecular masses in kDa are indicated on the left. For experimental details, see [5]. In later experiments, with better preservation of TG2 activity, MCA was replaced by 80% ethyleneglycol [8]. (B) Purified hRBC TG2 appears to be preponderantly in the GDP-bound form in the hydrodynamically compact, fast-moving electrophoretic conformation. Mobility shifts of the purified protein (lane 1) from slow to fast forms were examined upon mixing with GMP (lane 2), GDP (lane 3), and GTP (lane 4) by nondenaturing electrophoresis (in 3% agarose). For details, see [9].
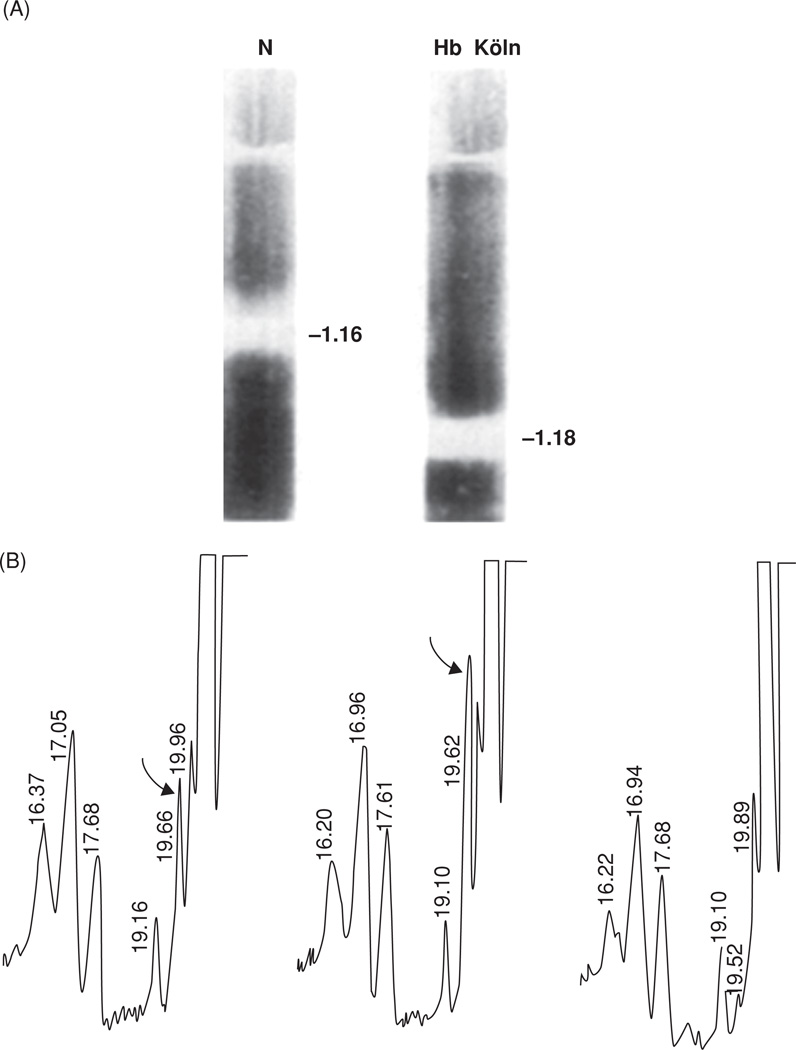
(A) Increased density of the red cell membranes from a patient with Hb Koln disease (right) compared with that of a normal individual (left), measured by sucrose density centrifugation. Density values, marked in g/mL, were computed from the refractive indices of the solutions at the points of highest turbidities of cell membranes. For details, see [18]. (B) Skeletal proteins of the Hb Koln erythrocyte membrane are modified by Nε (γ-glutaminyl)lysine side chain bridges, footprints of the activity of intracellular TG2. High-pressure liquid chromatography of the total proteolytic digest of SDS–DTT-insoluble membrane proteins reveals the presence of the isopeptide (left panel). The same is shown in the middle panel but with synthetic Nε (γ-glutaminyl)lysine peptide added. The right-hand panel corresponds to the first panel on the left, following treatment by γ-glutamylaminecyclotransferase, an enzyme specific for cleaving the isopeptide. Positions of Nε (γ-glutaminyl)lysine are marked by curved arrows. For details, see [18].
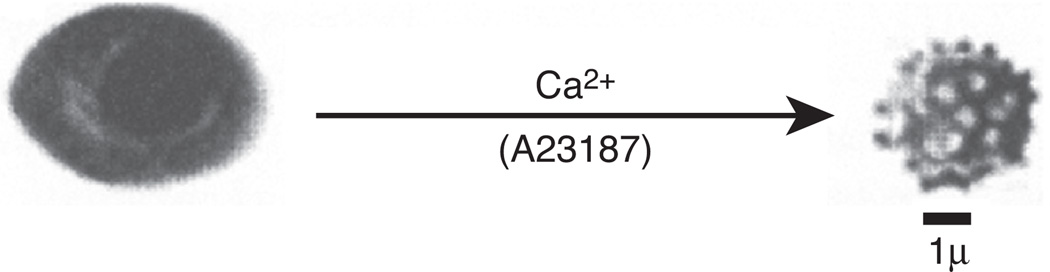
Change of shape in human red cells from discocyte to spheroechinocyte, brought about by Ca2+ overload, is illustrated by scanning electron micrography. The process of shape change is reversible only if the intracellular Ca2+ is removed within a short period of time; however, it becomes irreversible by longer exposure to the cation. Fixation of the abnormal spheroechinocytic shape—seen in some erythrocyte diseases—is due to the cross-linking of membrane skeletal proteins by Nε (γ-glutaminyl)lysine bonds under the catalytic influence of TG2. Inhibitors of the enzymatic reaction prevent loss of membrane plasticity, which otherwise would irreversibly freeze the shape of the red cell. For details, see [15].
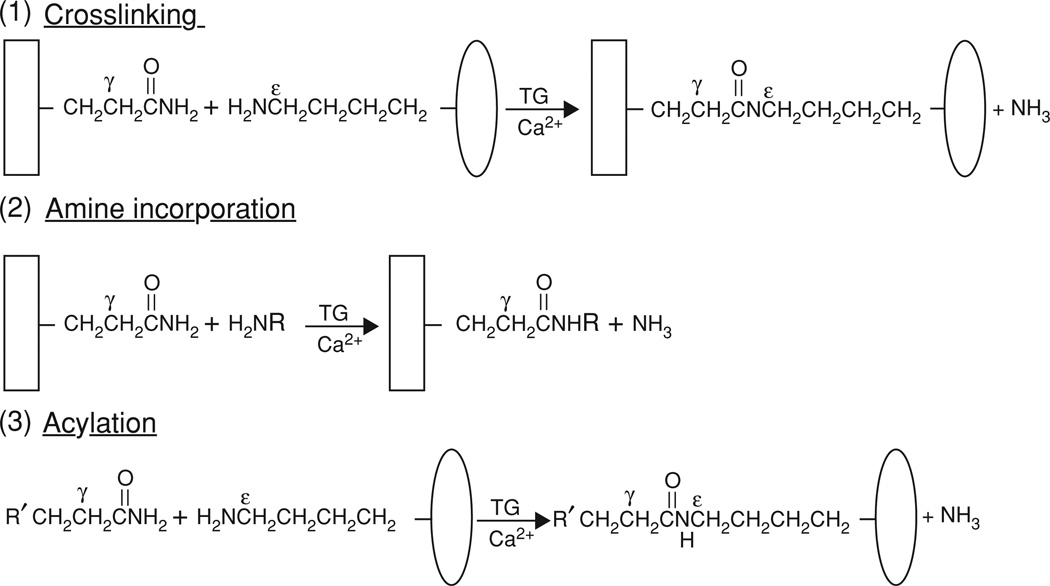
Reactions catalyzed by hRBC TG2 and other transglutaminases. Cross-linking of two proteins by Nε (γ-glutaminyl)lysine bonds (reaction 1) can be inhibited by the other two competing reactions shown. Incorporation of small primary amines (reaction 2) blocks the TG2-reactive γ-glutamyl cross-linking sites in the acceptor protein, whereas incorporation of small peptides with TG2-reactive glutaminyl residues blocks the ε-lysyl cross-linking sites of donor proteins (reaction 3). Labeled amines and glutamyl peptides are widely used for identifying and exploring potential protein substrates of transglutaminases; for a review, see [23].
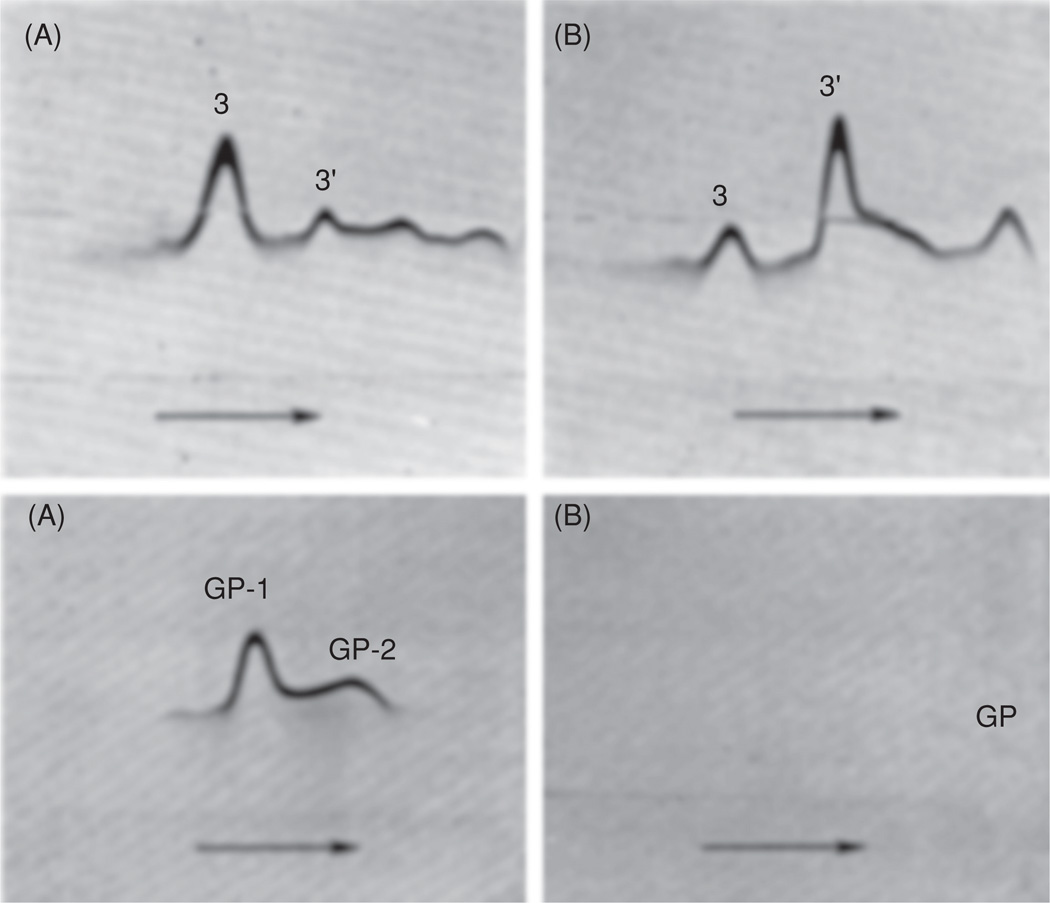
Pepstatin inhibits the proteolysis of anion transporter band 3 (top panels A and B) and glycophorin (bottom panels A and B), triggered in freshly isolated human erythrocytes by the influx of Ca2+. Red cells were incubated (12 h, 37°C) in Ca2+ (1.5 mM) and ionophore A23187 (10 µM) in the presence (A) and the absence (B) of pepstatin (2 mM). Membranes were isolated; proteins were extracted with SDS–DTT and electrophoresed in polyacrylamide gels horizontally (see arrows). After removal of SDS, cross-immunoelectrophoresis was performed in agarose (in the perpendicular direction) into which monospecific antibodies against the band 3 protein (top panels) or glycophorins (bottom panels) were incorporated. Immunoprecipitates corresponding to the intact anion transporter are marked as 3 and to its main breakdown product as 3′, whereas the intact glycophorins are denoted GP-1 and GP-2. The barely recognizable, small glycophorin fragments remaining in the absence of pepstatin are marked GP. For further details, see [25].
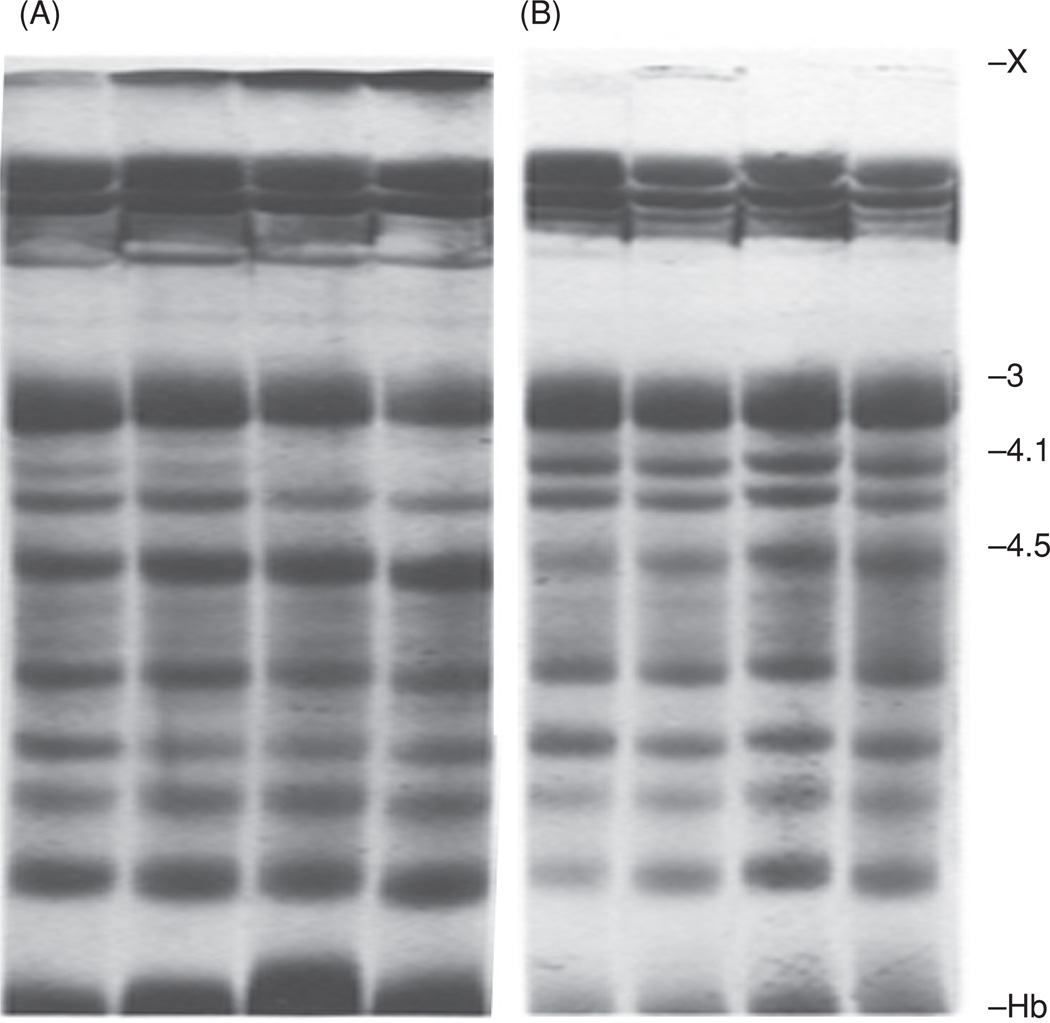
A direct, noncompetitive inhibitor of TG2, 2-[3(diallylamino)propionyl] benzothiophene, or DAPBT, effectively blocks the cross-linking of membrane skeletal proteins brought about by this enzyme when activated in the human red cell by an overload of Ca2+. The inhibitor also hinders proteolytic degradation of the band 3 anion transporter. Cells were incubated at 37°C with 2-mM Ca2+ and 20-µM ionophore A23187 for 1, 2, 4 and 8 h (corresponding to four gel profiles in each set from left to right) in the presence of 0.1 mM (set A) or 0.6 mM DAPBT (set B). The higher concentration of the inhibitor is seen to have prevented the formation of high-molecular-weight membrane protein polymers, marked X on top of the SDS-PAGE profiles, whereas lower concentration of DAPBT still allowed polymer production to proceed (albeit at a much slower rate than it would have without the presence of the compound). In SDS-PAGE tests, a sensitive indicator of blocking the cross-linking reaction by TG2 is preservation of staining intensity of the 4.1 band, as in (B). This panel also shows, that—at a concentration of 0.6 mM—DAPBT hindered the proteolytic degradation of the anion transporter band 3 into its main fragment, appearing at the position of band 4.5 (which corresponds to that of 3’ in cross-immunoelectrophoresis in Figure 5). For details, see [30].
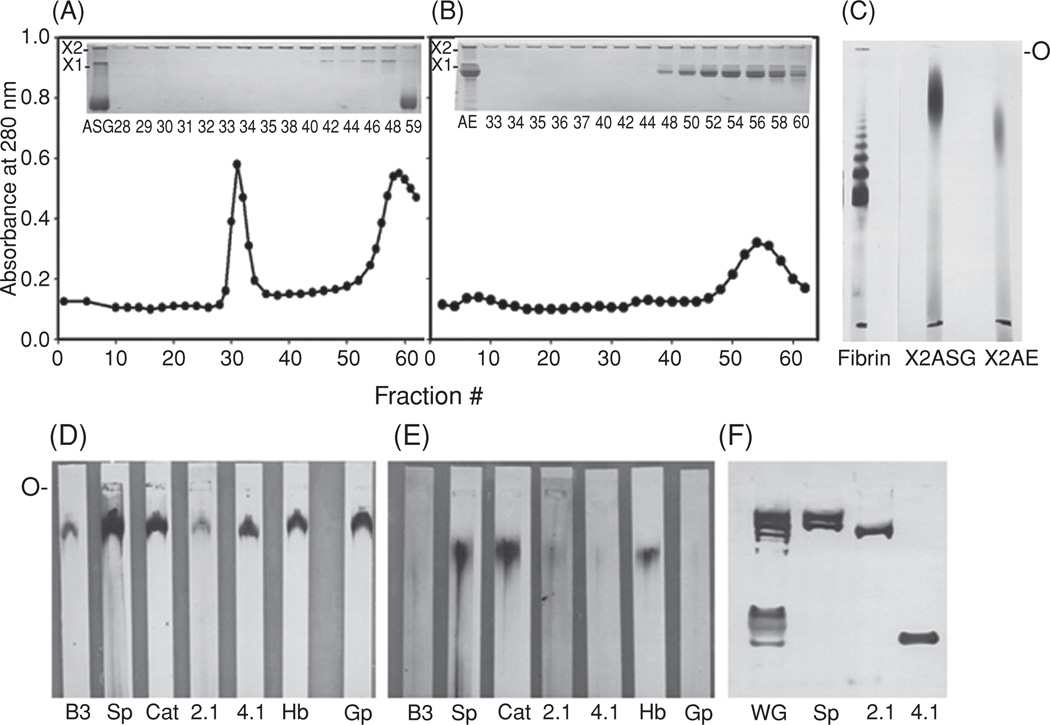
Properties of high-molecular-weight polymers isolated from Ca2+-treated human erythrocytes. Red cells were exposed to Ca2+ plus ionophore for 3 h; after hypotonic lysis, membranes were harvested and treated with 0.1 N NaOH. Proteins of the alkali-stripped ghosts (ASG) and alkali extract (AE) were fractionated by chromatography on Sepharose CL-4B; see (A) and (B). Insets to the panels show the SDS-PAGE profiles for the ASG and AE preparations applied to the column and for the fractions that contained X1 and X2 polymers, visualized on top of the separating and stacking gels. The X2 fractions from the ASG (A; fractions #29–35) and from the AE materials (B; fractions #33–40) were pooled, concentrated, and examined by SDS-agarose electrophoresis (C; O indicates origin). A nonreduced fibrin preparation, cross-linked by factor XIIIa, was employed as a molecular weight marker [32]. (D) and (E) are immunoblots of the purified X2 polymers of the alkali-stripped ghosts and from the alkali extract, following SDS electrophoresis on agarose (O marks origin), as in (C). Nitrocellulose blots were probed with various dilutions of antibodies, i.e., antisera to band 3 or B3 (1:300), spectrins or Sp (1:80,000), ankyrin or 2.1 (1:800), protein 4.1 or 4.1 (1:700), hemoglobin or Hb (1:13,000), glycophorin or Gp (1:7000), and an IgG to catalase or Cat (1:800). (F) A rabbit IgG raised against the cross-linked polymers from Ca2+-treated human erythrocytes [31], in 1:5000 dilution, reacted with several monomeric membrane skeletal protein on the SDS-PAGE profile (5% acrylamide) of whole ghosts of normal red cells (WG), which co-migrated with purified spectrins (Sp), ankyrin (2.1), and protein 4.1 (4.1), each of which was also recognized by the antibody. Bands on the immunoblot of lane WG above the position of 4.1 probably correspond to adducin and the anion transporter band 3 protein.
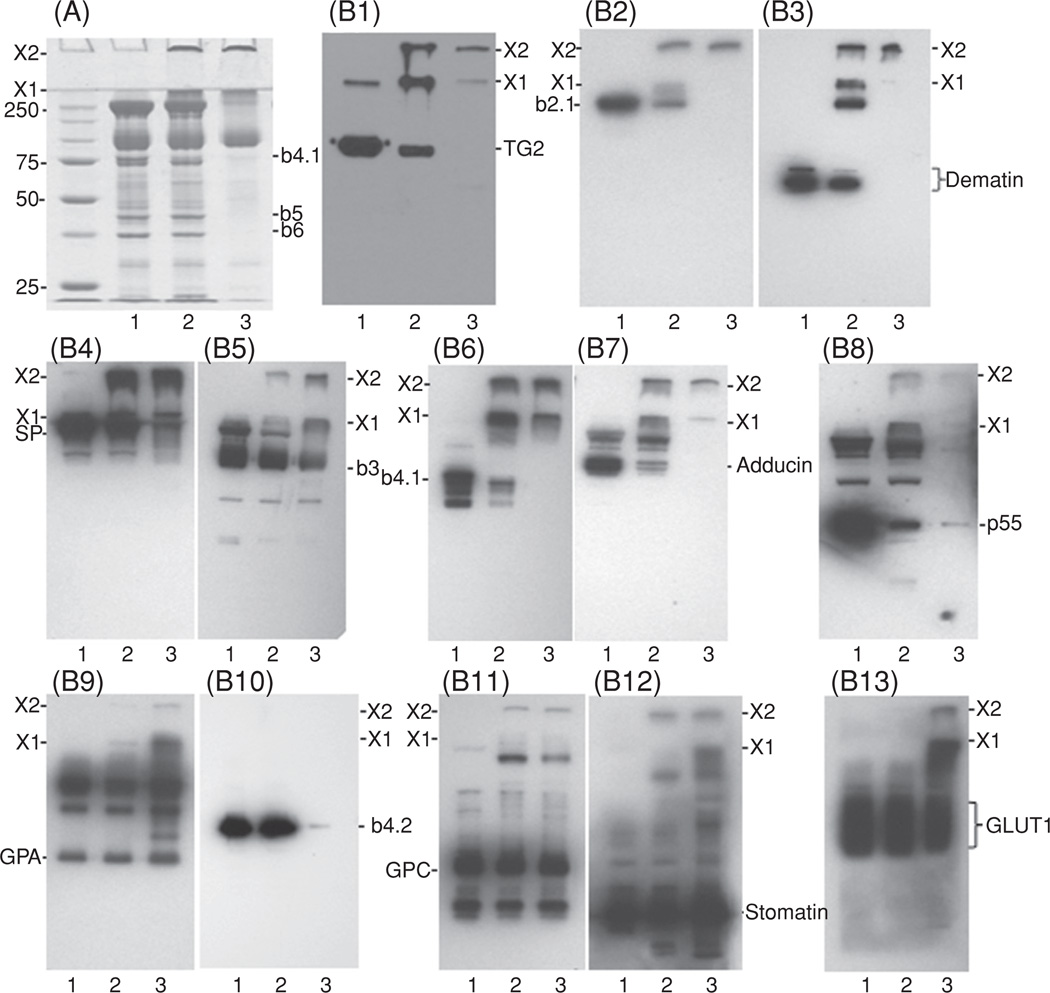
Cross-linked polymers, formed in human erythrocytes upon elevation of the concentration of internal Ca2+, are recognized by several antibodies from our new antibody repertoire. The polymers on the SDS-PAGE protein profiles of membrane proteins at the top of separating and stacking gels are marked as X1 and X2. Part (A) shows the Coomassie blue-stained gels of whole ghosts of control erythrocytes (lane 1), those with Ca2+ overload (lane 2), and of the alkali-stripped membranes of the latter (lane 3); molecular weight marker values are given in kDa; the position of actin is marked as band 5 or b5 and that of glyceraldehyde-3-phosphate dehydrogenase as band 6 or b6. PVDF transblots of the gels were probed with various dilutions of antibodies targeting transglutaminase 2 (TG2, B1; 1:20000), ankyrin (b2.1, B2; 1:20,000), dematin (B3; 1:5000), spectrins (SP, B4; 1:10,000), band 3 (b3, B5; 1:20,000), band 4.1 (b4.1, B6; 1:4000), adducin (B7; 1:3000), p55 (B8; 1:4000), glycophorin A (GPA, B9; 1:1000), band 4.2 (b4.2, B10; 1:200,000), glycophorin C (GPC, B11; 1:1000), stomatin (B12; 1:100), and glucose transporter 1 (GLUT1, B13; 1:20,000). (See insert for color representation.)
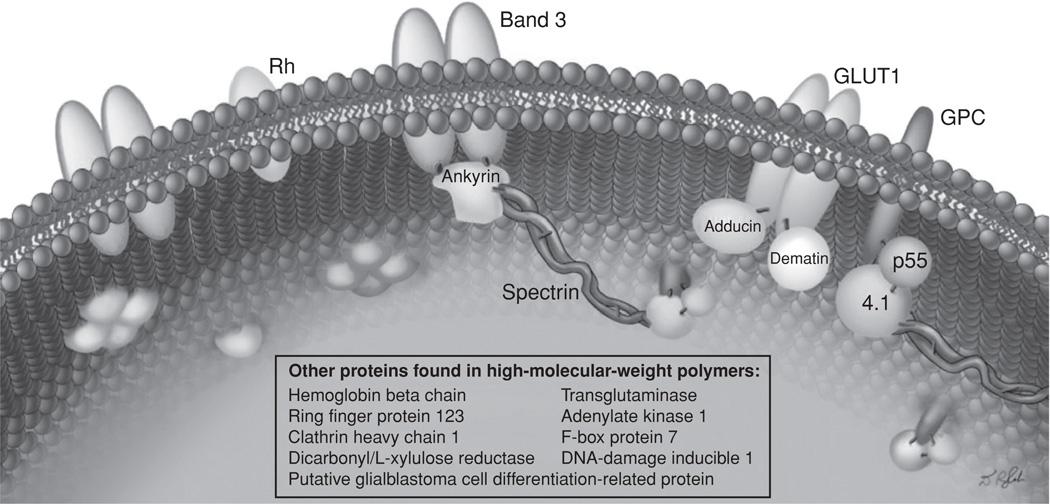
Illustration of a section of the inner surface of alkali-stripped ghosts from Ca2+-loaded erythrocytes indicating scattered clusters of protein polymers, covalently linked to transmembrane proteins or to p55, which—through its palmitoyl moiety—is partially embedded in the lipid bi-layer. The Nε (γ-glutamyl)lysine side chain bridges between constituent polypeptide chains of the polymer are shown by red lines. Without remodeling of the membrane skeleton (as in the left portion of the picture), only the cytoplasmic domains of proteins such as band 3, GLUT1, the Rhesus blood group CcEe antigen, and p55 are seen. (See insert for color representation.)
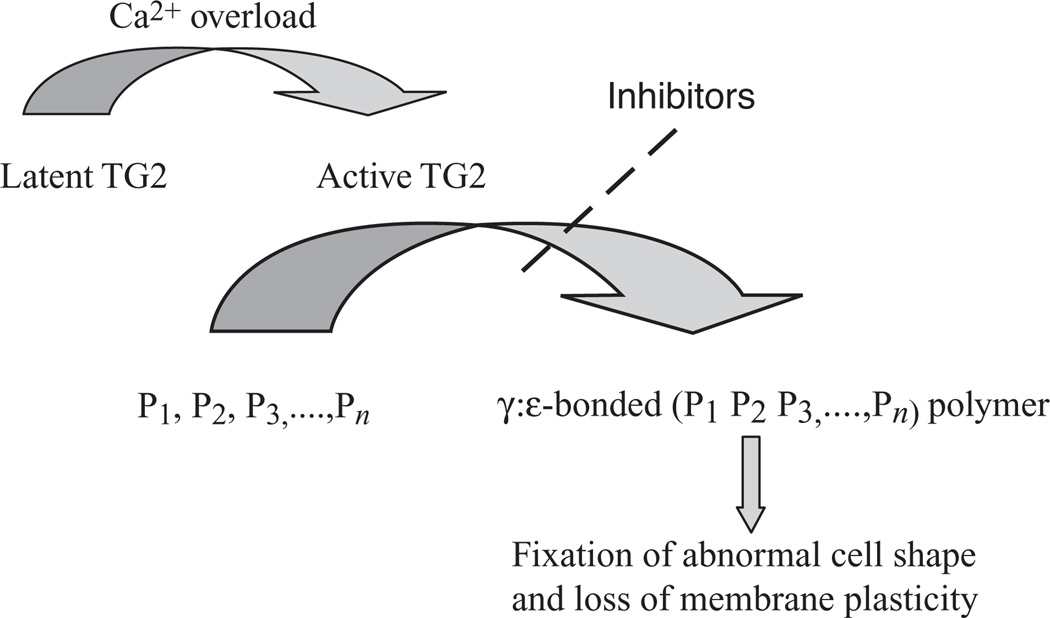
The Ca2+-triggered, transglutaminase-mediated protein crosslinking cascade in cells.
Similar articles
-
Tissue transglutaminase (TG2) facilitates phosphatidylserine exposure and calpain activity in calcium-induced death of erythrocytes.Cell Death Differ. 2007 Oct;14(10):1842-4. doi: 10.1038/sj.cdd.4402193. Epub 2007 Jul 6. Cell Death Differ. 2007. PMID: 17612588 Free PMC article. No abstract available.
-
Tissue transglutaminase and its role in human cancer progression.Adv Enzymol Relat Areas Mol Biol. 2011;78:247-93. doi: 10.1002/9781118105771.ch6. Adv Enzymol Relat Areas Mol Biol. 2011. PMID: 22220476 Review. No abstract available.
-
Transglutaminase 2 at the crossroads between cell death and survival.Adv Enzymol Relat Areas Mol Biol. 2011;78:197-246. doi: 10.1002/9781118105771.ch5. Adv Enzymol Relat Areas Mol Biol. 2011. PMID: 22220475 Review. No abstract available.
-
Physiopathological roles of human transglutaminase 2.Adv Enzymol Relat Areas Mol Biol. 2011;78:47-95. doi: 10.1002/9781118105771.ch2. Adv Enzymol Relat Areas Mol Biol. 2011. PMID: 22220472 Review. No abstract available.
-
Osmotic resistance of high-density erythrocytes in transglutaminase 2-deficient mice.Biochem Biophys Res Commun. 2002 Mar 15;291(5):1123-7. doi: 10.1006/bbrc.2002.6558. Biochem Biophys Res Commun. 2002. PMID: 11883932
Cited by
-
Calcium in red blood cells-a perilous balance.Int J Mol Sci. 2013 May 8;14(5):9848-72. doi: 10.3390/ijms14059848. Int J Mol Sci. 2013. PMID: 23698771 Free PMC article. Review.
-
Measurement of posttransfusion red cell survival with the biotin label.Transfus Med Rev. 2014 Jul;28(3):114-25. doi: 10.1016/j.tmrv.2014.03.003. Epub 2014 Apr 5. Transfus Med Rev. 2014. PMID: 24969019 Free PMC article. Review.
References
-
- Turner PM, Lorand L. Complexation of fibronectin with tissue transglutaminase. Biochemistry. 1989;28:628–635. - PubMed
-
- LeMosy EK, Erickson HP, Beyer WF, Jr., Radek JT, Jeong JM, Murthy SN, Lorand L. Visualization of purified fibronectin-transglutaminase complexes. J. Biol. Chem. 1992;267:7880–7885. - PubMed
-
- Hang J, Zemskov EA, Lorand L, Belkin AM. Identification of a novel recognition sequence for fibronectin within the NH2-terminal beta-sandwich domain of tissue transglutaminase. J. Biol. Chem. 2005;280:23675–23683. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical