

The Cbln family of proteins interact with multiple signaling pathways
- PMID: 22220752
- PMCID: PMC3342465
- DOI: 10.1111/j.1471-4159.2012.07648.x
The Cbln family of proteins interact with multiple signaling pathways
Abstract
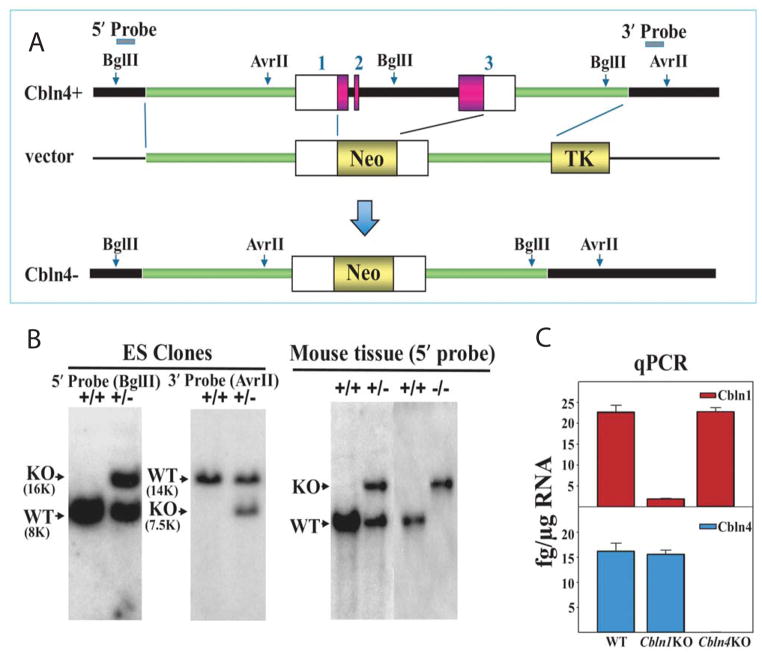
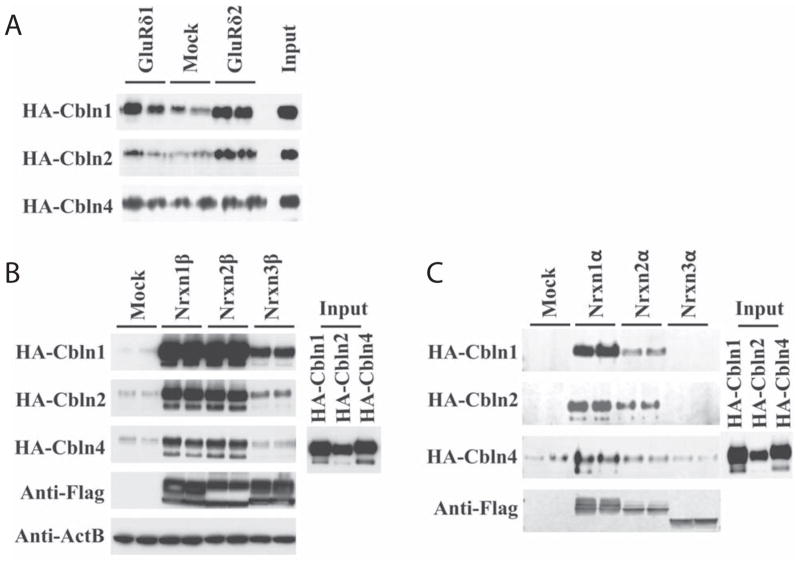
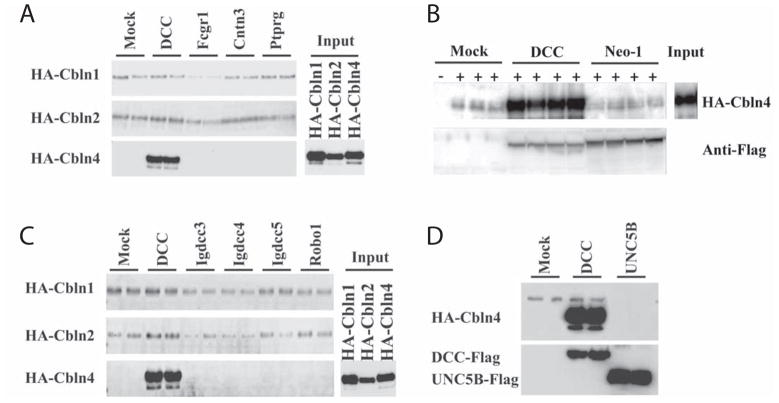
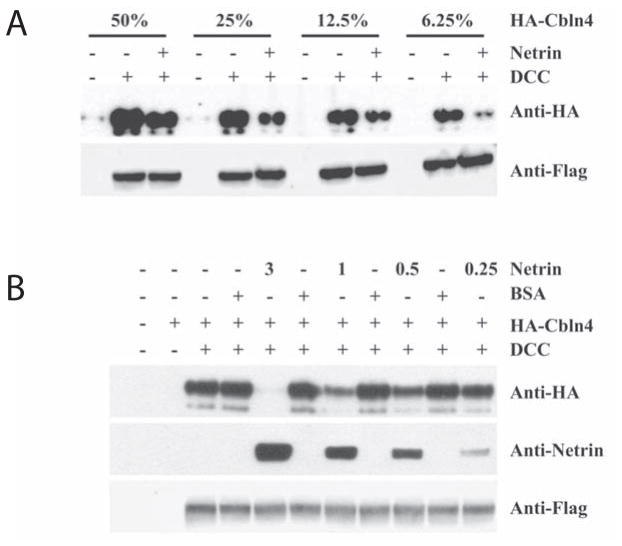
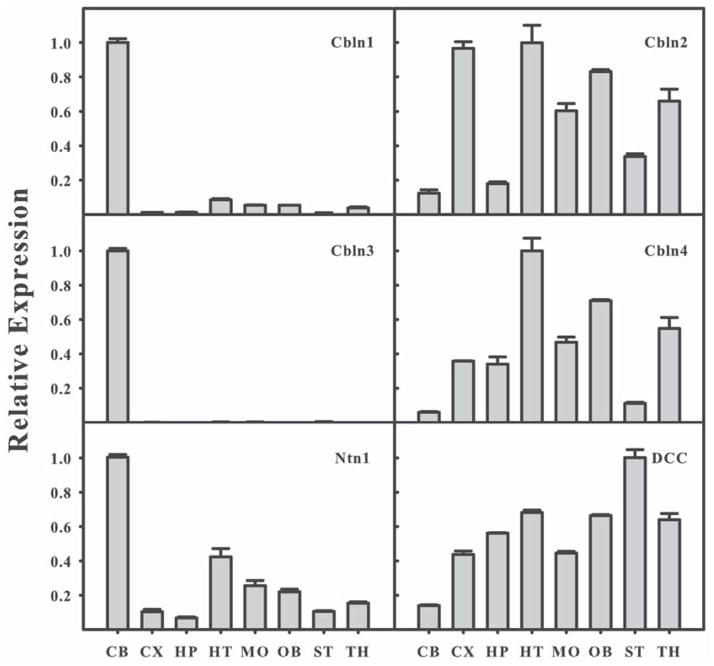
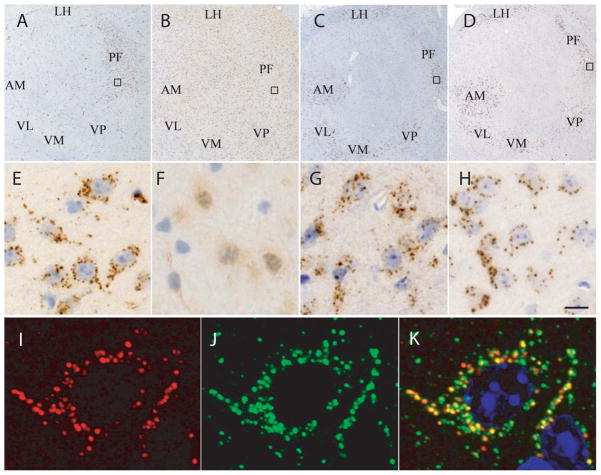
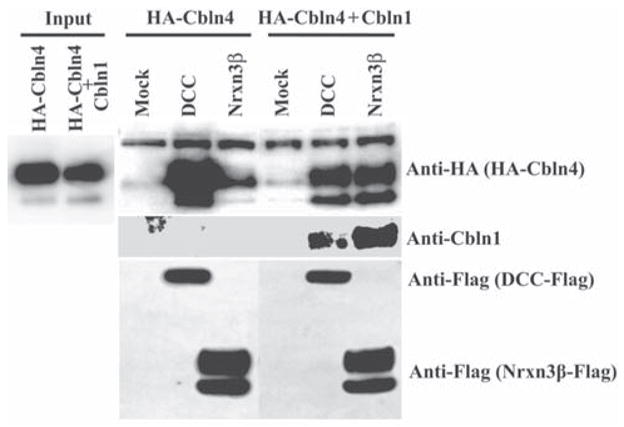
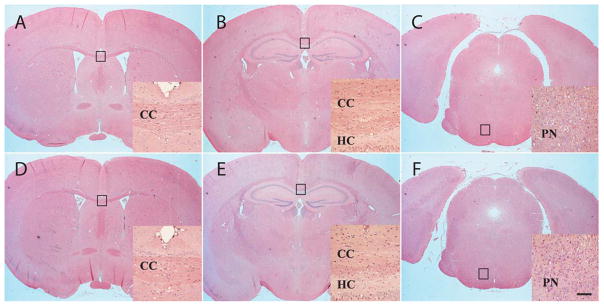
Cerebellin precursor protein (Cbln1) is essential for synapse integrity in cerebellum through assembly into complexes that bridge pre-synaptic β-neurexins (Nrxn) to post-synaptic GluRδ2. However, GluRδ2 is largely cerebellum-specific, yet Cbln1 and its little studied family members, Cbln2 and Cbln4, are expressed throughout brain. Therefore, we investigated whether additional proteins mediate Cbln family actions. Whereas Cbln1 and Cbln2 bound to GluRδ2 and Nrxns1-3, Cbln4 bound weakly or not at all, suggesting it has distinct binding partners. In a candidate receptor-screening assay, Cbln4 (but not Cbln1 or Cbln2) bound selectively to the netrin receptor, (deleted in colorectal cancer (DCC) in a netrin-displaceable fashion. To determine whether Cbln4 had a netrin-like function, Cbln4-null mice were generated. Cbln4-null mice did not phenocopy netrin-null mice. Cbln1 and Cbln4 were likely co-localized in neurons thought to be responsible for synaptic changes in striatum of Cbln1-null mice. Furthermore, complexes containing Cbln1 and Cbln4 had greatly reduced affinity to DCC but increased affinity to Nrxns, suggesting a functional interaction. However, Cbln4-null mice lacked the striatal synaptic changes seen in Cbln null mice. Thus, Cbln family members interact with multiple receptors/signaling pathways in a subunit composition-dependent manner and have independent functions with Cbln4 potentially involved in the less well-characterized role of netrin/DCC in adult brain.
© 2012 The Authors. Journal of Neurochemistry © 2012 International Society for Neurochemistry.
Conflict of interest statement
None of the authors have any financial interests or conflicts of interest in the data presented in the manuscript.
Figures
Comment in
-
Cerebellins: capstones to bridge the synaptic cleft.J Neurochem. 2012 Jun;121(5):697-9. doi: 10.1111/j.1471-4159.2012.07675.x. J Neurochem. 2012. PMID: 22288876 No abstract available.
References
-
- Ackerman SL, Kozak LP, Przyborski SA, Rund LA, Boyer BB, Knowles BB. The mouse rostral cerebellar malformation gene encodes an UNC-5-like protein. Nature. 1997;386:838–842. - PubMed
-
- Araki K, Meguro H, Kushiya E, Takayama C, Inoue Y, Mishina M. Selective expression of the glutamate receptor channel delta 2 subunit in cerebellar Purkinje cells. Biochem Biophys Res Commun. 1993;197:1267–1276. - PubMed
-
- Bao D, Pang Z, Morgan JI. The structure and proteolytic processing of Cbln1 complexes. J Neurochem. 2005;95:618–629. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
