

Ketamine-induced neuroapoptosis in the fetal and neonatal rhesus macaque brain
- PMID: 22222480
- PMCID: PMC3433282
- DOI: 10.1097/ALN.0b013e318242b2cd
Ketamine-induced neuroapoptosis in the fetal and neonatal rhesus macaque brain
Abstract
Background: Exposure of rhesus macaque fetuses for 24 h or neonates for 9 h to ketamine anesthesia causes neuroapoptosis in the developing brain. The current study clarifies the minimum exposure required for and the extent and spatial distribution of ketamine-induced neuroapoptosis in rhesus fetuses and neonates.
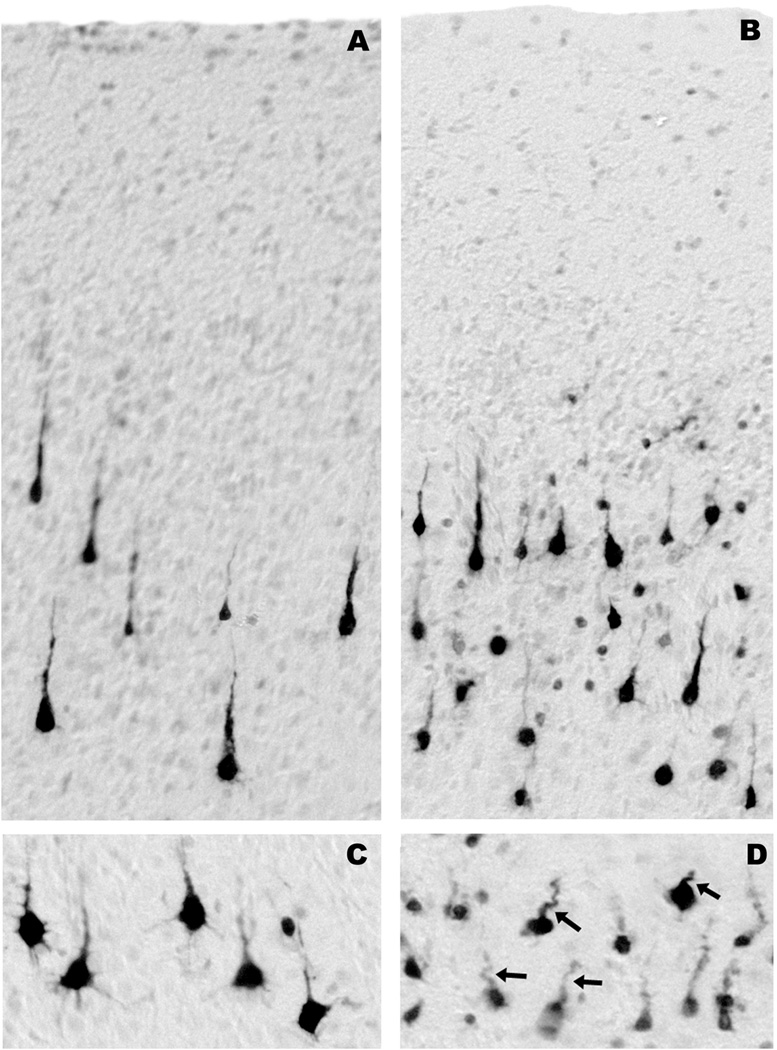
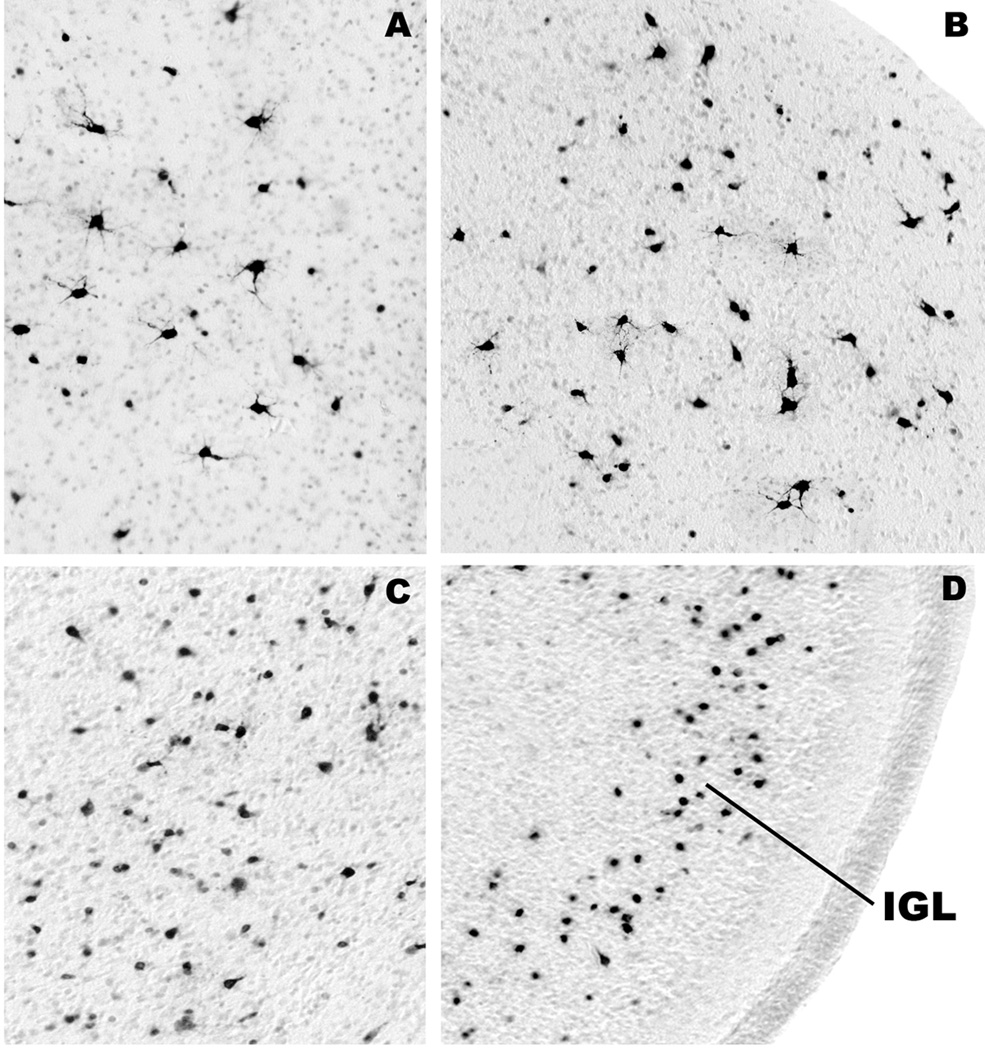
Method: Ketamine was administered by IV infusion for 5 h to postnatal day 6 rhesus neonates or to pregnant rhesus females at 120 days' gestation (full term = 165 days). Three hours later, fetuses were delivered by cesarean section, and the fetal and neonatal brains were studied for evidence of apoptotic neurodegeneration, as determined by activated caspase-3 staining.
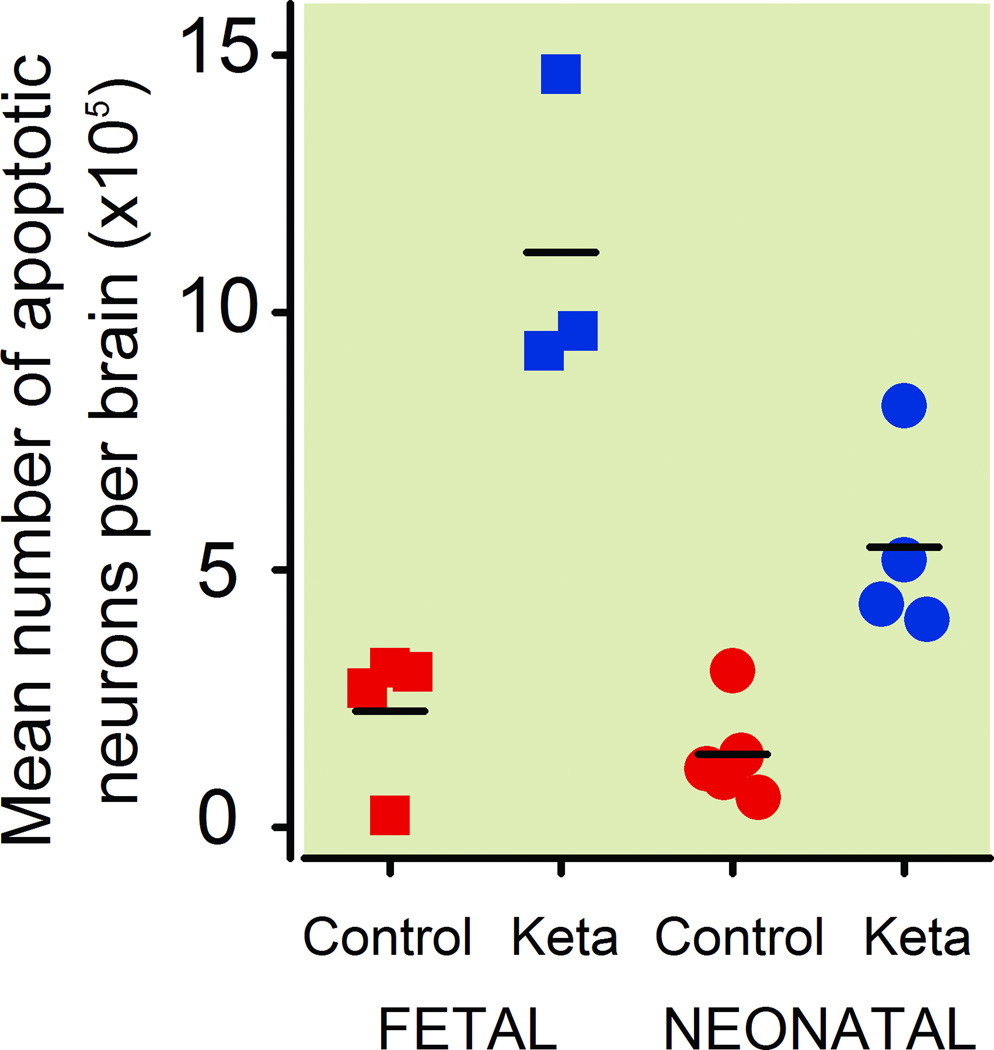
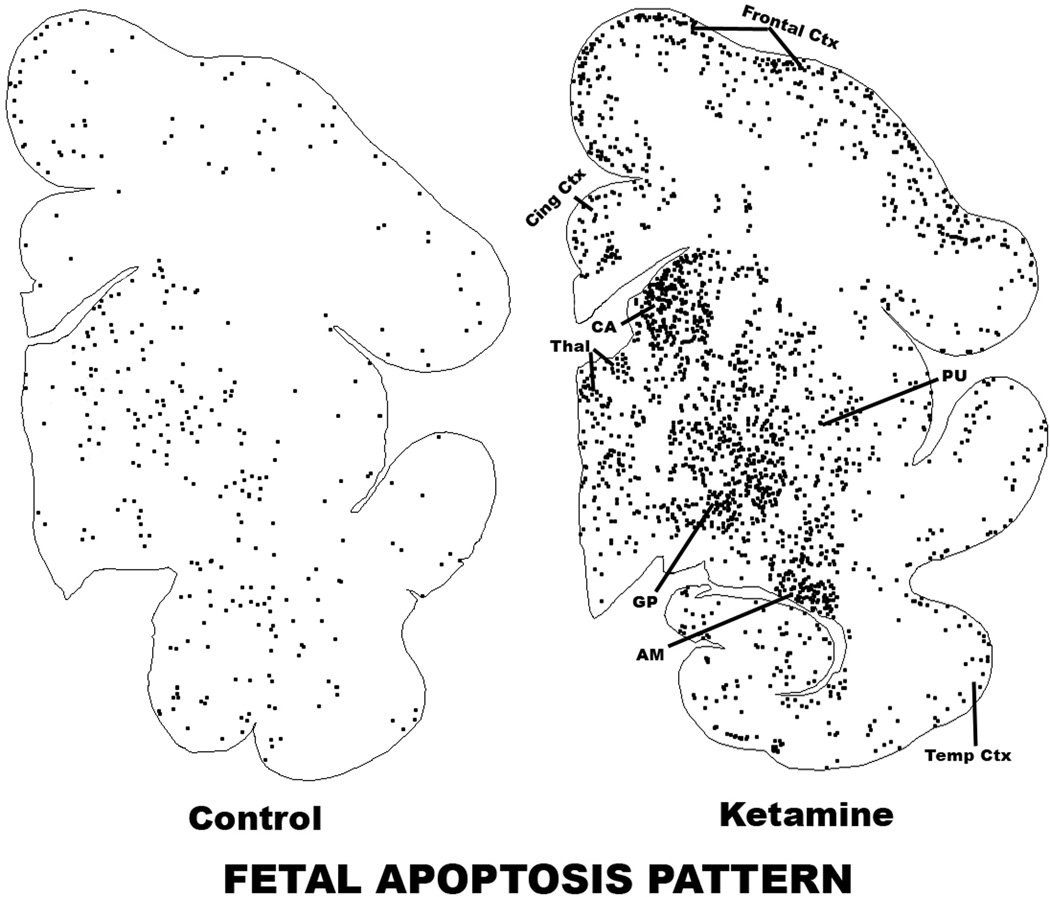
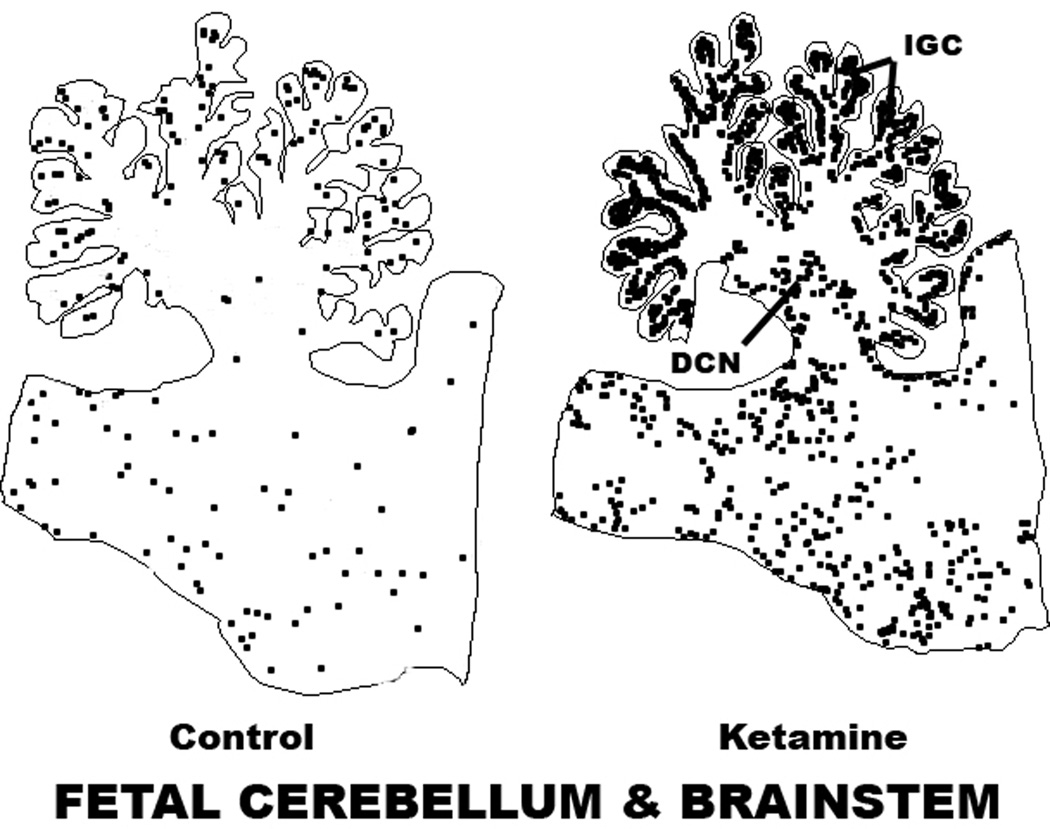
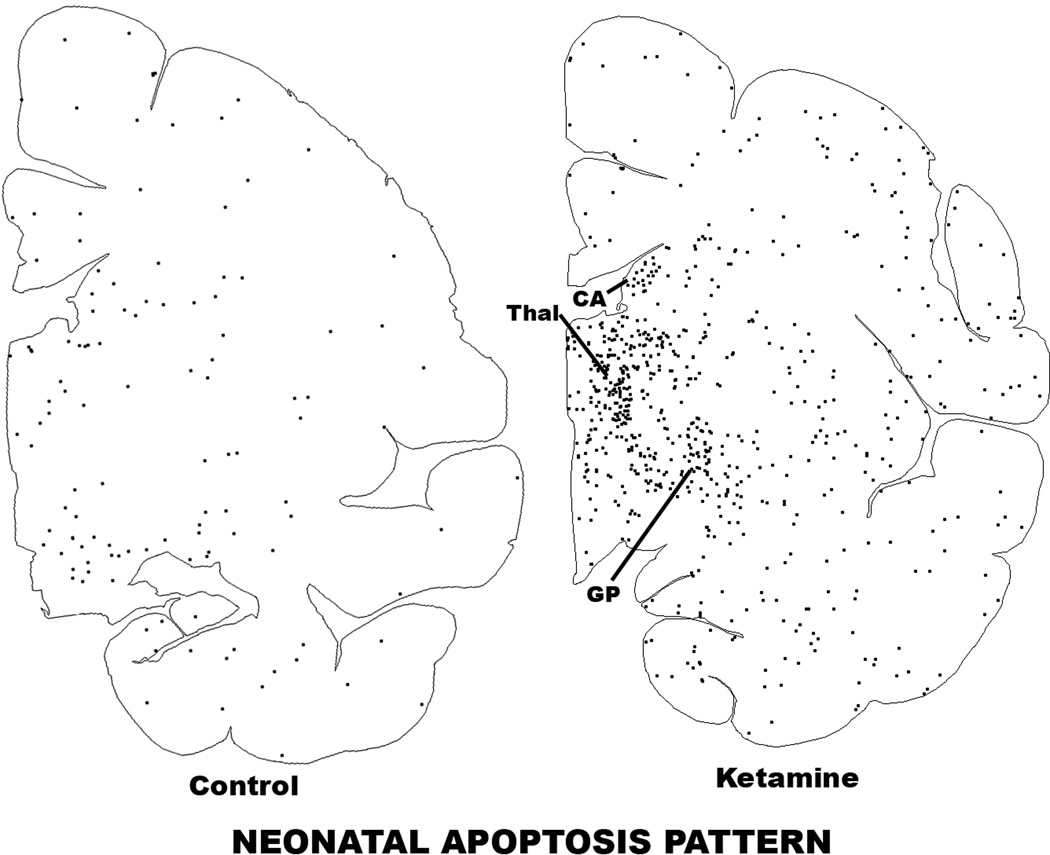
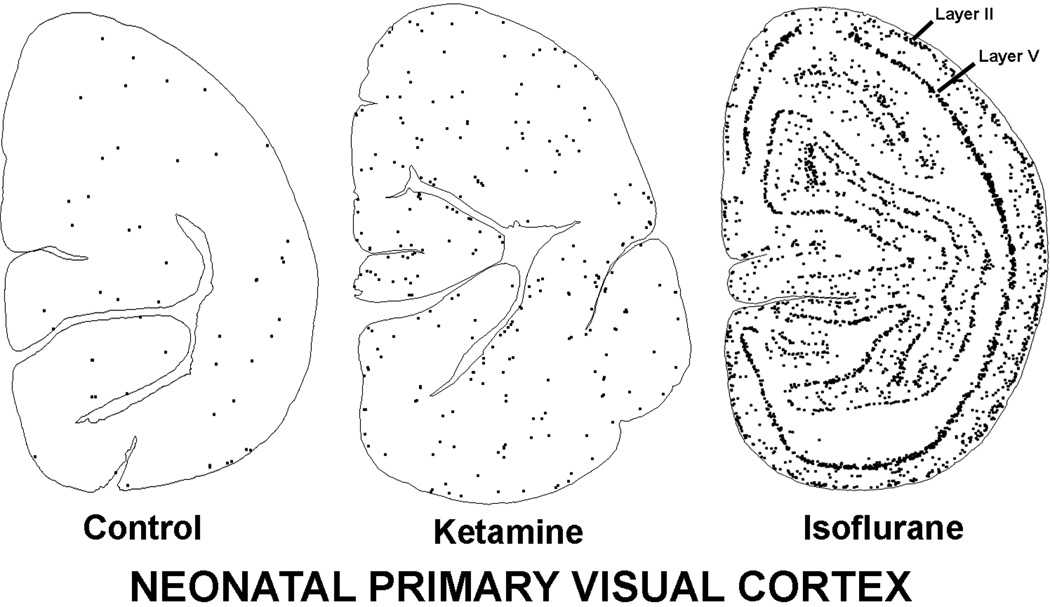
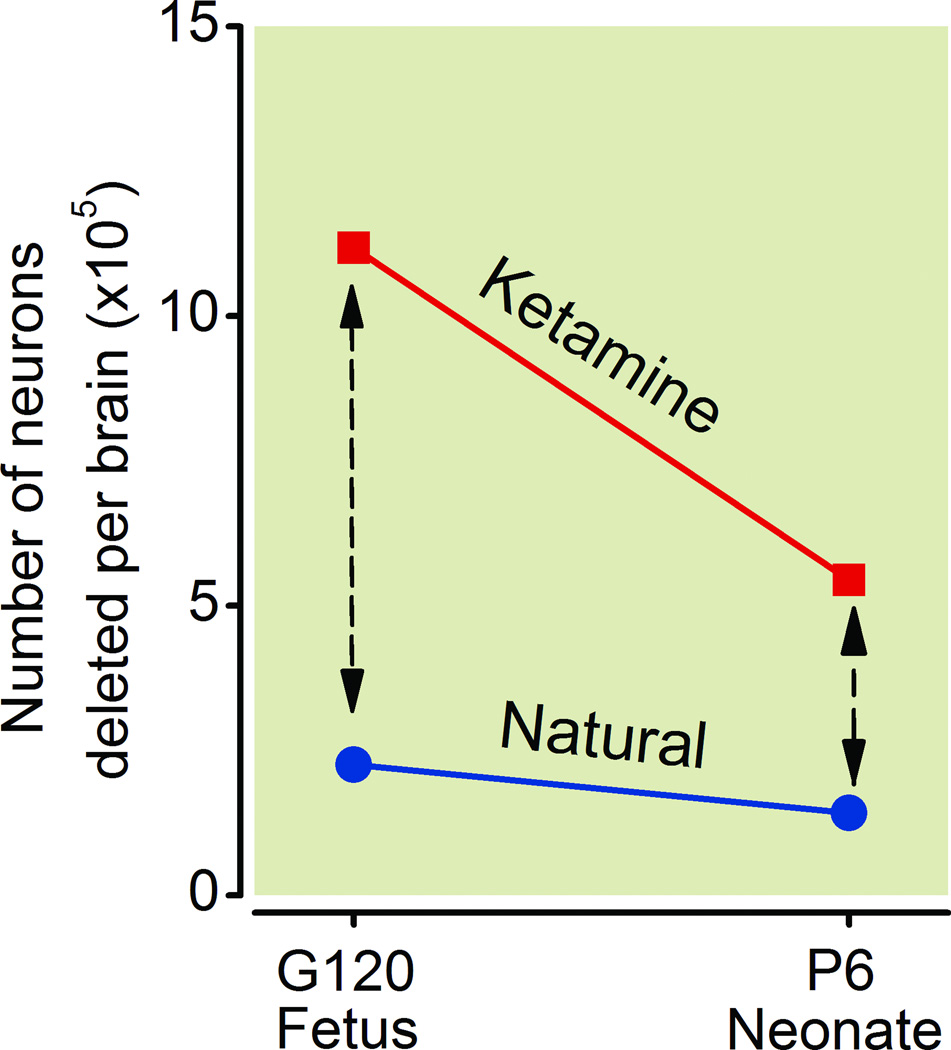
Results: Both the fetal (n = 3) and neonatal (n = 4) ketamine-exposed brains had a significant increase in apoptotic profiles compared with drug-naive controls (fetal n = 4; neonatal n = 5). Loss of neurons attributable to ketamine exposure was 2.2 times greater in fetuses than in neonates. The pattern of neurodegeneration in fetuses was different from that in neonates, and all subjects exposed at either age had a pattern characteristic for that age.
Conclusion: The developing rhesus macaque brain is sensitive to the apoptogenic action of ketamine at both a fetal and neonatal age, and exposure duration of 5 h is sufficient to induce a significant neuroapoptosis response at either age. The pattern of neurodegeneration induced by ketamine in fetuses was different from that in neonates, and loss of neurons attributable to ketamine exposure was 2.2 times greater in the fetal than neonatal brains.
Figures
References
-
- Ikonomidou C, Bosch F, Miksa M, Bittigau P, Vöckler J, Dikranian K, Tenkova T, Stevoska V, Turski L, Olney JW. Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain. Science. 1999;283:70–74. - PubMed
-
- Ikonomidou C, Bittigau P, Ishimaru MJ, Wozniak DF, Koch C, Genz K, Price MT, Stefovska V, Hörster F, Tenkova T, Dikranian K, Olney JW. Ethanol-induced apoptotic neurodegeneration and fetal alcohol syndrome. Science. 2000;287:1056–1060. - PubMed
-
- Olney JW, Tenkova T, Dikranian K, Qin YQ, Labruyere J, Ikonomidou C. Ethanol-induced apoptotic neurodegeneration in the developing C57BL/6 mouse brain. Dev Brain Res. 2002;133:115–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
