

The induction of a nucleoplasmic reticulum by prelamin A accumulation requires CTP:phosphocholine cytidylyltransferase-α
- PMID: 22223883
- PMCID: PMC3258109
- DOI: 10.1242/jcs.091009
The induction of a nucleoplasmic reticulum by prelamin A accumulation requires CTP:phosphocholine cytidylyltransferase-α
Abstract
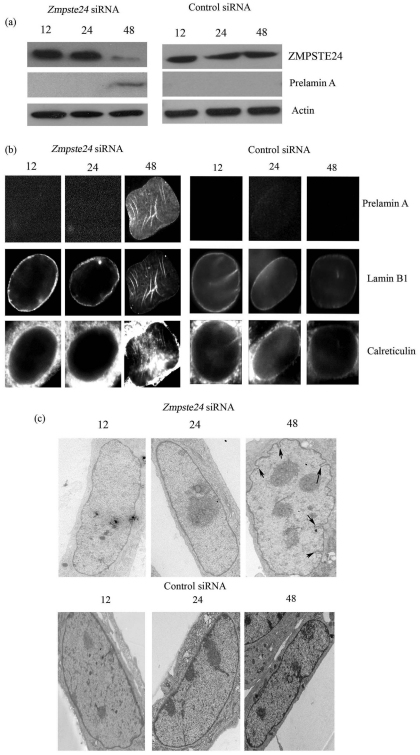
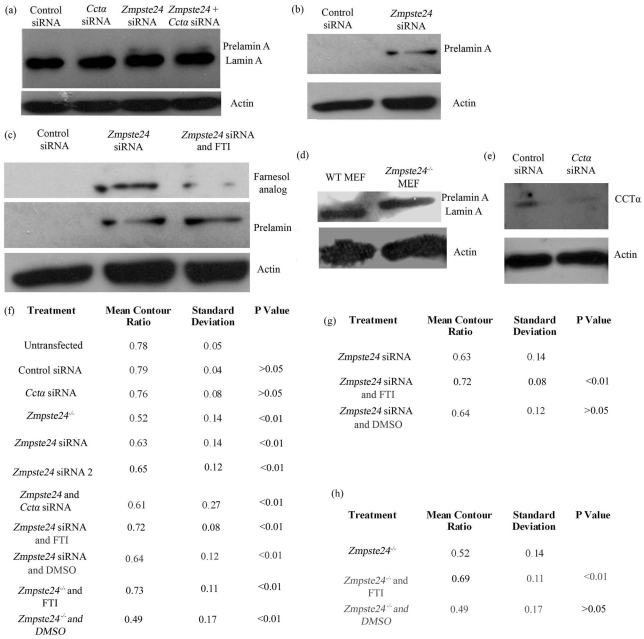
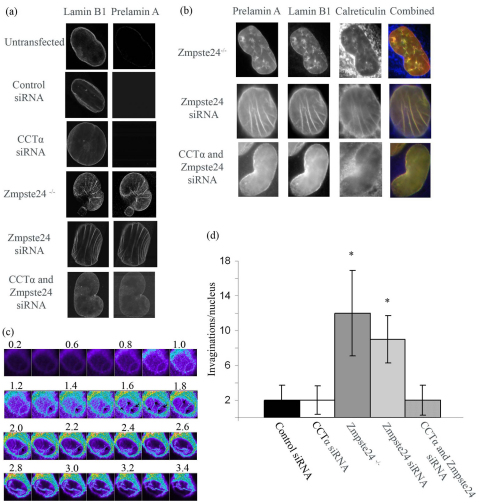
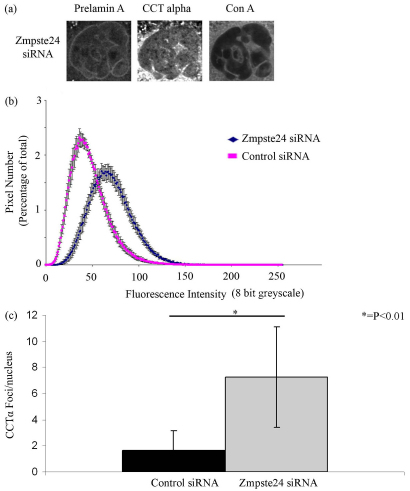
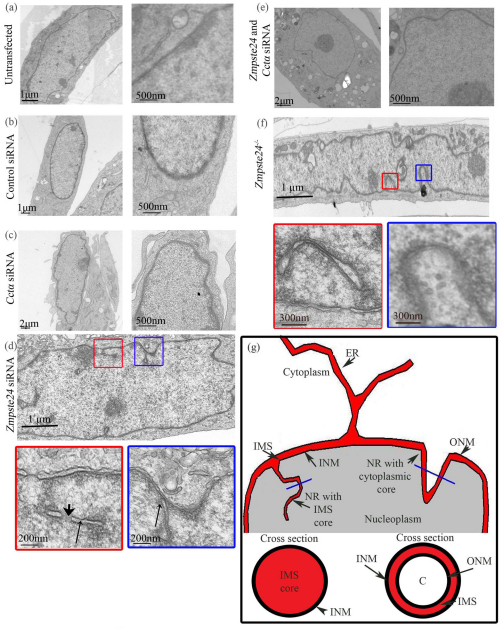
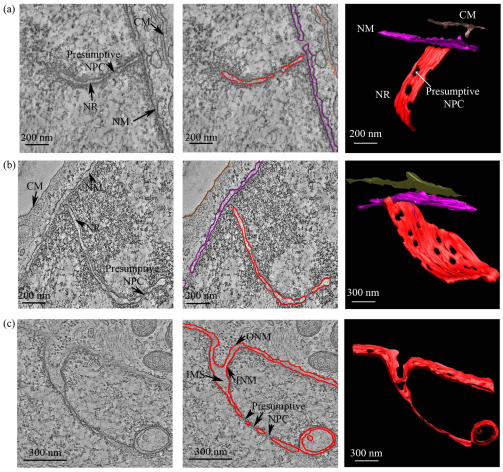
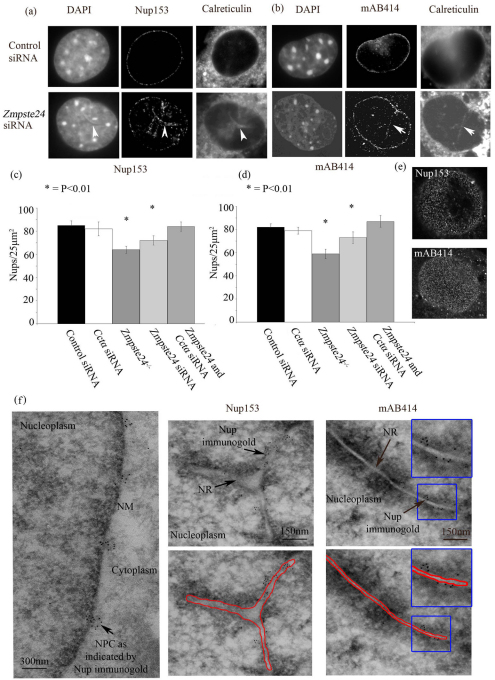
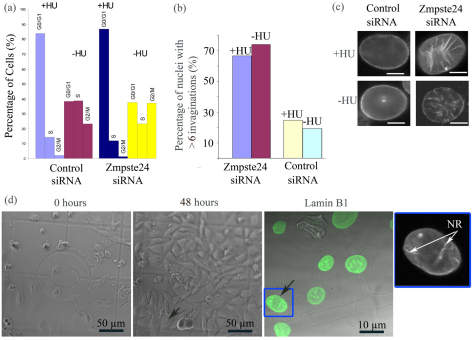
Farnesylated prelamin A accumulates when the final endoproteolytic maturation of the protein fails to occur and causes a dysmorphic nuclear phenotype; however, the morphology and mechanisms of biogenesis of these changes remain unclear. We show here that acute prelamin A accumulation after reduction in the activity of the ZMPSTE24 endoprotease by short interfering RNA knockdown, results in the generation of a complex nucleoplasmic reticulum that depends for its formation on the enzyme CTP:phosphocholine-cytidylyltransferase-α (CCT-α, also known as choline-phosphate cytidylyltransferase A). This structure can form during interphase, confirming that it is independent of mitosis and therefore not a consequence of disordered nuclear envelope assembly. Serial-section dual-axis electron tomography reveals that these invaginations can take two forms: one in which the inner nuclear membrane infolds alone with an inter membrane space interior, and the other in which an invagination of both nuclear membranes occurs, enclosing a cytoplasmic core. Both types of invagination can co-exist in one nucleus and both are frequently studded with nuclear pore complexes (NPC), which reduces NPC abundance on the nuclear surface.
Figures
Similar articles
-
Expansion of the nucleoplasmic reticulum requires the coordinated activity of lamins and CTP:phosphocholine cytidylyltransferase alpha.Mol Biol Cell. 2008 Jan;19(1):237-47. doi: 10.1091/mbc.e07-02-0179. Epub 2007 Oct 24. Mol Biol Cell. 2008. PMID: 17959832 Free PMC article.
-
CTP:phosphocholine cytidylyltransferase α (CCTα) and lamins alter nuclear membrane structure without affecting phosphatidylcholine synthesis.Biochim Biophys Acta. 2011 Jun;1811(6):377-85. doi: 10.1016/j.bbalip.2011.04.001. Epub 2011 Apr 9. Biochim Biophys Acta. 2011. PMID: 21504799
-
Analysis of prelamin A biogenesis reveals the nucleus to be a CaaX processing compartment.Mol Biol Cell. 2008 Dec;19(12):5398-408. doi: 10.1091/mbc.e08-07-0704. Epub 2008 Oct 15. Mol Biol Cell. 2008. PMID: 18923140 Free PMC article.
-
Prelamin A farnesylation and progeroid syndromes.J Biol Chem. 2006 Dec 29;281(52):39741-5. doi: 10.1074/jbc.R600033200. Epub 2006 Nov 7. J Biol Chem. 2006. PMID: 17090536 Review.
-
Prelamin A-mediated nuclear envelope dynamics in normal and laminopathic cells.Biochem Soc Trans. 2011 Dec;39(6):1698-704. doi: 10.1042/BST20110657. Biochem Soc Trans. 2011. PMID: 22103510 Review.
Cited by
-
Nuclear Morphological Remodeling in Human Granulocytes Is Linked to Prenylation Independently from Cytoskeleton.Cells. 2020 Nov 20;9(11):2509. doi: 10.3390/cells9112509. Cells. 2020. PMID: 33233551 Free PMC article.
-
Condensins exert force on chromatin-nuclear envelope tethers to mediate nucleoplasmic reticulum formation in Drosophila melanogaster.G3 (Bethesda). 2014 Dec 30;5(3):341-52. doi: 10.1534/g3.114.015685. G3 (Bethesda). 2014. PMID: 25552604 Free PMC article.
-
Highlight on the dynamic organization of the nucleus.Nucleus. 2017 Jan 2;8(1):2-10. doi: 10.1080/19491034.2016.1243634. Epub 2016 Oct 7. Nucleus. 2017. PMID: 27715428 Free PMC article. Review.
-
Nuclear shape changes are induced by knockdown of the SWI/SNF ATPase BRG1 and are independent of cytoskeletal connections.PLoS One. 2013;8(2):e55628. doi: 10.1371/journal.pone.0055628. Epub 2013 Feb 6. PLoS One. 2013. PMID: 23405182 Free PMC article.
-
Formation of a nucleoplasmic reticulum requires de novo assembly of nascent phospholipids and shows preferential incorporation of nascent lamins.Sci Rep. 2017 Aug 7;7(1):7454. doi: 10.1038/s41598-017-07614-w. Sci Rep. 2017. PMID: 28785031 Free PMC article.
References
-
- Bkaily G., Avedanian L., Jacques D. (2009). Nuclear membrane receptors and channels as targets for drug development in cardiovascular diseases. Can. J. Physiol. Pharmacol. 87, 108-119 - PubMed
-
- Bootman M. D., Fearnley C., Smyrnias I., MacDonald F., Roderick H. L. (2009). An update on nuclear calcium signalling. J. Cell Sci. 122, 2337-2350 - PubMed
-
- Busch A., Kiel T., Heupel W. M., Wehnert M., Hubner S. (2009). Nuclear protein import is reduced in cells expressing nuclear envelopathy-causing lamin A mutants. Exp. Cell Res. 315, 2373-2385 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
