

Generation of chimeric rhesus monkeys
- PMID: 22225614
- PMCID: PMC3264685
- DOI: 10.1016/j.cell.2011.12.007
Generation of chimeric rhesus monkeys
Abstract
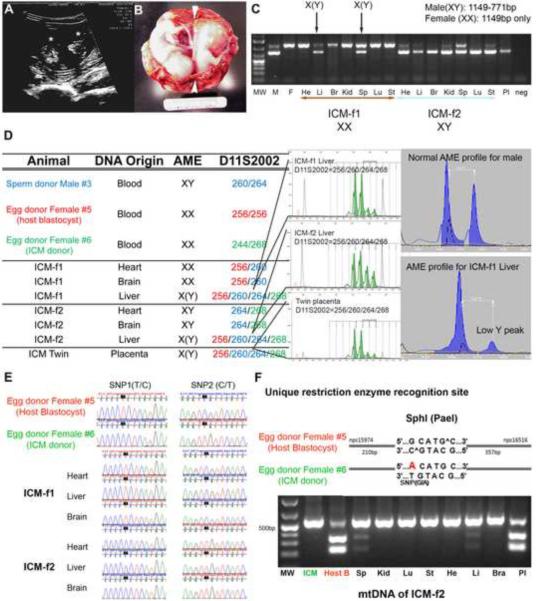
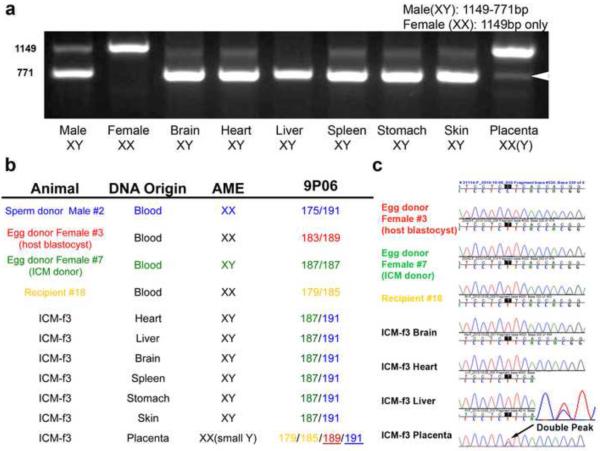
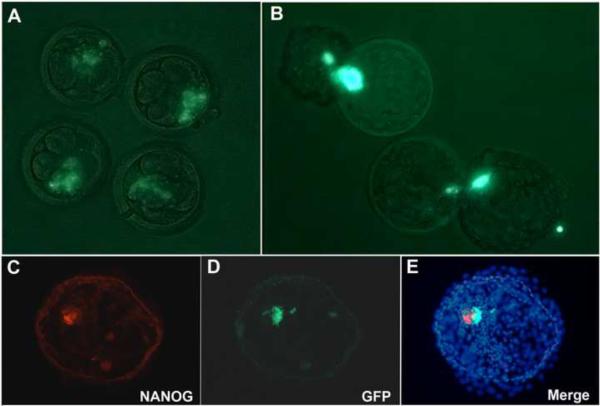
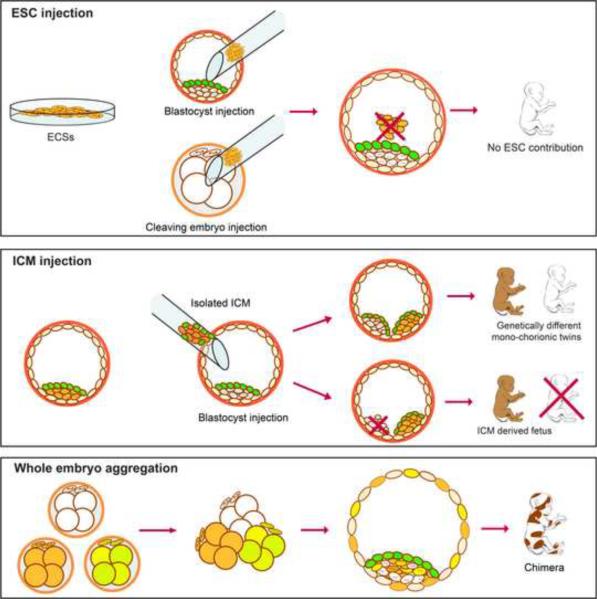
Totipotent cells in early embryos are progenitors of all stem cells and are capable of developing into a whole organism, including extraembryonic tissues such as placenta. Pluripotent cells in the inner cell mass (ICM) are the descendants of totipotent cells and can differentiate into any cell type of a body except extraembryonic tissues. The ability to contribute to chimeric animals upon reintroduction into host embryos is the key feature of murine totipotent and pluripotent cells. Here, we demonstrate that rhesus monkey embryonic stem cells (ESCs) and isolated ICMs fail to incorporate into host embryos and develop into chimeras. However, chimeric offspring were produced following aggregation of totipotent cells of the four-cell embryos. These results provide insights into the species-specific nature of primate embryos and suggest that a chimera assay using pluripotent cells may not be feasible.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

Comment in
-
Chimeric primates: embryonic stem cells need not apply.Cell. 2012 Jan 20;148(1-2):19-21. doi: 10.1016/j.cell.2011.12.016. Epub 2012 Jan 5. Cell. 2012. PMID: 22225613
References
-
- Beddington RS, Robertson EJ. An assessment of the developmental potential of embryonic stem cells in the midgestation mouse embryo. Development. 1989;105:733–737. - PubMed
-
- Bradley A, Evans M, Kaufman MH, Robertson E. Formation of germ-line chimaeras from embryo-derived teratocarcinoma cell lines. Nature. 1984;309:255–256. - PubMed
-
- Brem G, Tenhumberg H, Krausslich H. Chimerism in cattle through microsurgical aggregation of morulae. Theriogenology. 1984;22:609–613. - PubMed
-
- Brons IG, Smithers LE, Trotter MW, Rugg-Gunn P, Sun B, Chuva de Sousa Lopes SM, Howlett SK, Clarkson A, Ahrlund-Richter L, Pedersen RA, et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature. 2007;448:191–195. - PubMed
-
- Byrne JA, Pedersen DA, Clepper LL, Nelson M, Sanger WG, Gokhale S, Wolf DP, Mitalipov SM. Producing primate embryonic stem cells by somatic cell nuclear transfer. Nature. 2007;450:497–502. - PubMed
Publication types
MeSH terms
Grants and funding
- P51 RR000163/RR/NCRR NIH HHS/United States
- HD059946/HD/NICHD NIH HHS/United States
- HD063276/HD/NICHD NIH HHS/United States
- HD047675/HD/NICHD NIH HHS/United States
- HD018185/HD/NICHD NIH HHS/United States
- R01 HD057121/HD/NICHD NIH HHS/United States
- R01 HD059946/HD/NICHD NIH HHS/United States
- K01 RR000163/RR/NCRR NIH HHS/United States
- RR000163/RR/NCRR NIH HHS/United States
- P30 HD018185/HD/NICHD NIH HHS/United States
- U54 HD018185/HD/NICHD NIH HHS/United States
- P01 HD047675/HD/NICHD NIH HHS/United States
- R01 HD063276/HD/NICHD NIH HHS/United States
- HD057121/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
