

Roles of long and short replication initiation proteins in the fate of IncP-1 plasmids
- PMID: 22228734
- PMCID: PMC3294859
- DOI: 10.1128/JB.06395-11
Roles of long and short replication initiation proteins in the fate of IncP-1 plasmids
Abstract
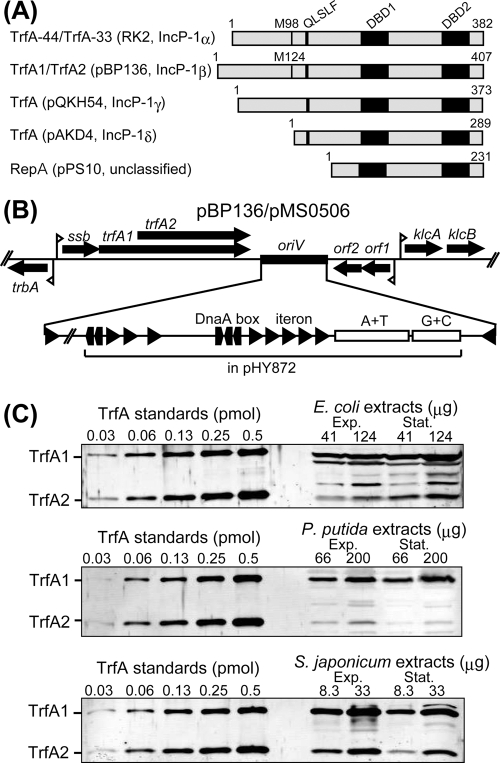
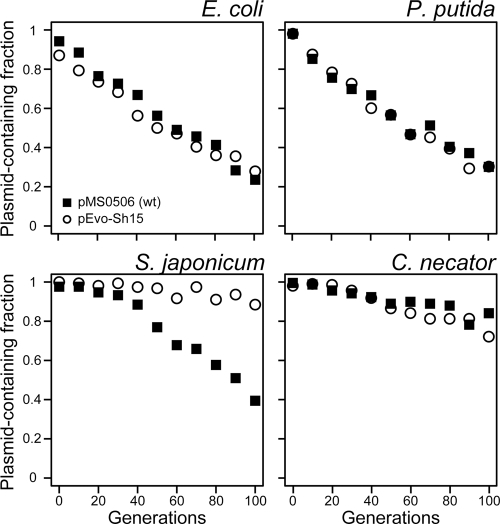
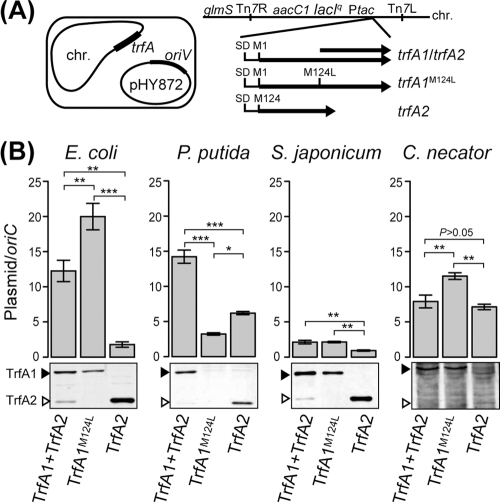
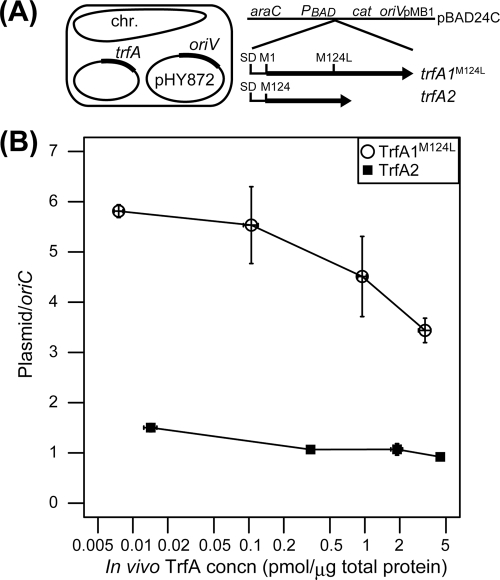
Broad-host-range IncP-1 plasmids generally encode two replication initiation proteins, TrfA1 and TrfA2. TrfA2 is produced from an internal translational start site within trfA1. While TrfA1 was previously shown to be essential for replication in Pseudomonas aeruginosa, its role in other bacteria within its broad host range has not been established. To address the role of TrfA1 and TrfA2 in other hosts, efficiency of transformation, plasmid copy number (PCN), and plasmid stability were first compared between a mini-IncP-1β plasmid and its trfA1 frameshift variant in four phylogenetically distant hosts: Escherichia coli, Pseudomonas putida, Sphingobium japonicum, and Cupriavidus necator. TrfA2 was sufficient for replication in these hosts, but the presence of TrfA1 enhanced transformation efficiency and PCN. However, TrfA1 did not contribute to, and even negatively affected, long-term plasmid persistence. When trfA genes were cloned under a constitutive promoter in the chromosomes of the four hosts, strains expressing either both TrfA1 and TrfA2 or TrfA1 alone, again, generally elicited a higher PCN of an IncP1-β replicon than strains expressing TrfA2 alone. When a single species of TrfA was produced at different concentrations in E. coli cells, TrfA1 maintained a 3- to 4-fold higher PCN than TrfA2 at the same TrfA concentrations, indicating that replication mediated by TrfA1 is more efficient than that by TrfA2. These results suggest that the broad-host-range properties of IncP-1 plasmids are essentially conferred by TrfA2 and the intact replication origin alone but that TrfA1 is nonetheless important to efficiently establish plasmid replication upon transfer into a broad range of hosts.
Figures
References
-
- Abhyankar MM, Reddy JM, Sharma R, Bullesbach E, Bastia D. 2004. Biochemical investigations of control of replication initiation of plasmid R6K. J. Biol. Chem. 279:6711–6719 - PubMed
-
- Adamczyk M, Jagura-Burdzy G. 2003. Spread and survival of promiscuous IncP-1 plasmids. Acta Biochim. Pol. 50:425–453 - PubMed
-
- Bagdasarian M, et al. 1981. Specific-purpose plasmid cloning vectors. II. Broad host range, high copy number, RSF1010-derived vectors, and a host-vector system for gene cloning in Pseudomonas. Gene 16:237–247 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
