

Review
doi: 10.1038/nrmicro2714.
Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes
Affiliations
- PMID: 22230951
- PMCID: PMC3313462
- DOI: 10.1038/nrmicro2714
Item in Clipboard
Review
Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes
Nat Rev Microbiol.
.
Abstract
In little more than 30 years, Lyme disease, which is caused by the spirochaete Borrelia burgdorferi, has risen from relative obscurity to become a global public health problem and a prototype of an emerging infection. During this period, there has been an extraordinary accumulation of knowledge on the phylogenetic diversity, molecular biology, genetics and host interactions of B. burgdorferi. In this Review, we integrate this large body of information into a cohesive picture of the molecular and cellular events that transpire as Lyme disease spirochaetes transit between their arthropod and vertebrate hosts during the enzootic cycle.
Figures
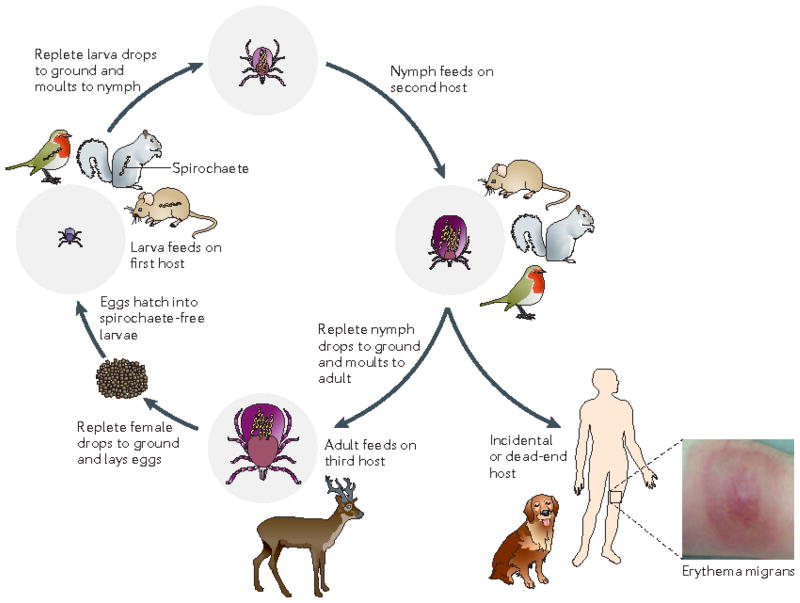
The enzootic cycle of Borrelia burgdorferi. Ixodes spp. ticks undergo a three-stage life cycle — larva, nymph and adult — with one blood meal per stage. Although some Borrelia spp. that cause relapsing fever can be passed from adult to egg (transovarial transmission), this does not occur with B. burgdorferi, so each generation of tick must acquire a B. burgdorferi infection anew. Larval ticks feed on many different animals, including Peromyscus spp. mice, squirrels and birds. B. burgdorferi infection is acquired by feeding on an infected reservoir animal, and the bacterium is retained during the subsequent stages (that is, trans-stadially) after each blood meal and moult,. Nymphs feed on a similar range of hosts to larvae; transmission of spirochaetes to a competent reservoir host by a feeding nymph perpetuates the enzootic cycle for the next generation of larval ticks. Although small mammals are usually thought of as the primary reservoirs for Lyme disease spirochaetes, studies have called attention to the importance of migratory birds as disseminators of spirochaetes over large distances,. Adult ticks are not generally important for maintenance of B. burgdorferi in the wild, as they feed predominantly on larger animals such as deer, which are incompetent hosts for B. burgdorferi. However, deer are important for maintenance of the tick population because adult ticks mate on them. Although all three stages of Ixodes scapularis can feed on humans, nymphs are responsible for the vast majority of spirochaete transmission to humans. It is unknown whether infected humans can transmit spirochaetes to feeding larvae, and humans are generally considered dead-end hosts and not part of the enzootic cycle. Dogs are probably incidental hosts and not part of the enzootic cycle.

a | Freeze-fracture electron micrographs showing the convex and concave leaflets of the inner and outer membranes (IM and OM, respectively). Integral membrane proteins (particles) are considerably more abundant in the IM; the density of OM particles is much lower than that in a prototypical Gram-negative bacterium. Scale bars represent 500 nm. b | Cryoelectron tomograms of the ends of borrelial cells showing IM, OM, peptidoglycan (PG), flagellar motor and filament assemblies (FMFA), chemoreceptor arrays (CR) and an external layer comprising outer-surface lipoproteins (Osps). Upper scale bar represents 1 μm, lower scale bar represents 100 nm. c | Cryoelectron tomographic cross-section showing the IM, OM, periplasmic space (PS) and periplasmic flagella (PFs). Scale bar represents 50 nm. d | Longitudinal cryoelectron tomographic slice showing a ribbon of nine flagellar filaments wrapping around the IM in a right-handed helix. Scale bar represents 200 nm. Part a is reproduced, with permission, from REF. c (1994) American Society for Microbiology (ASM). Part b is reproduced, with permission, from REF. c (2009) ASM, and from REF. c (2011) ASM. Parts c,d are reproduced, with permission, from REF. c (2009) ASM.
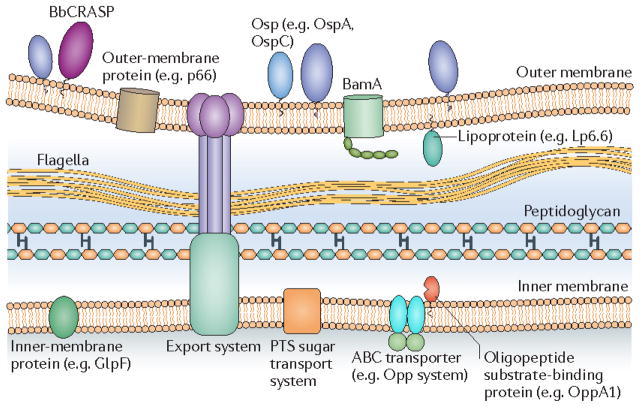
This schematic of the borrelial cell envelope shows the outer membrane, flagellar filaments, peptidoglycan, and cytoplasmic inner membrane. The outer membrane contains outer-surface lipoproteins (Osps) in high density and β-barrel outer-membrane-spanning proteins such as BamA in low density. The inner membrane is rich in integral membrane proteins, many of which are transporters. BbCRASP, complement regulator-acquiring protein; OppA1, oligopeptide permease A1; PTS, phosphotransferase system.

a | The histidine kinase 1 (Hk1)–response regulatory protein 1 (Rrp1) and alternative RNA polymerase σ-factor RpoS global regulatory systems. Binding of unidentified ligands to the periplasmic sensor domains (D1 and D2) of the hybrid histidine kinase Hk1 initiates a phosphorelay that activates the diguanylyl cyclase activity of Rrp1, resulting in the production of cyclic di-GMP (c-di-GMP)–,. Phosphodiesterase A (PdeA) and PdeB degrade c-di-GMP to 5′-phosphoguanylyl-(3′– 5′)-guanosine (pGpG) and GMP, respectively,. Activation of Rrp2 in vitro and in vivo occurs via the high-energy phosphoryl donor acetyl-phosphate rather than by its presumptive cognate histidine kinase, Hk2 (REF. 48). The function of Hk2 is currently unknown. Phosphorylated Rrp2, Borrelia oxidative stress regulator (BosR) and RpoN initiate transcription of rpoS,,–. This is depicted as a trimeric complex, but the precise interactions between these proteins have yet to be determined. Putative BosR-binding sites (BSs) containing the direct repeat sequence TAAATTAAAT are shown; −24/−12 is the RpoN-binding site in the rpoS promoter. RpoS in turn induces the expression of genes that are required during the mammalian-host phase of the spirochaete life cycle and represses the expression of tick-phase genes.b | Expression of the Hk1–Rrp1 and RpoS global regulatory systems during the B. burgdorferi life cycle,,–,,. In the flat nymph, both the Hk1–Rrp1 and the Rrp2–RpoN–RpoS systems are inactive and only tick-phase genes are expressed. The nymphal blood meal activates both the Hk1 Rrp1 and Rrp2–RpoN–RpoS pathways. Expression of mammalian-phase genes begins in concert with downregulation of tick-phase genes. Following inoculation into a mammalian host, the spirochaetes complete the process of adaptation; the Hk1–Rrp1 pathway is inactive, the Rrp2–RpoN–RpoS pathway is active, mammalian-phase genes are expressed and tick-phase genes are repressed. During larval acquisition of spirochaetes, Hk1–Rrp1 is activated, probably at the feeding site, whereas the Rrp2–RpoN–RpoS system is inactivated. Mammalian-phase genes are repressed, expression of tick-phase genes begins and ingested spirochaetes bind to the larval midgut epithelium via OspA and possibly other receptors–. GGDEF, a conserved motif present in diguanylyl cyclases; Hpt, histidine-containing phosphotransfer domain; HTH, helix–turn–helix doain; N, amino; PAS, putative sensor domain for Hk2; Rec, receiver domain.
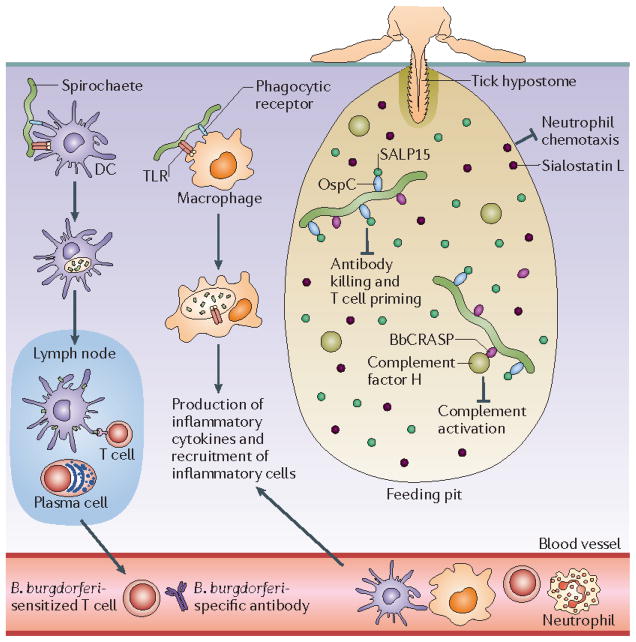
The tick creates a feeding pit with its mouthparts, using its hypostome (a barbed protuberance) as an anchor to the skin of its host. Initial salivary secretions form a cement cone around the hypostome that further anchors the tick during feeding. Subsequently, the tick produces copious amounts of saliva containing a plethora of bioactive agents that Borrelia burgdorferi exploits to help establish infection. The diagram shows just a few of these bioactive agents, including SALP15 (which binds to the spirochaetes and inhibits killing of the bacteria and T cell priming) and sialostatin L (which blocks neutrophil chemotaxis),. A group of borrelial surface lipoproteins, collectively referred to as BbCRASPs (complement regulator-acquiring surface proteins), bind complement factor H, preventing activation of the alternative complement pathway –. B. burgdorferi cells are recognized by innate immune effector cells such as dendritic cells (DCs), neutrophils and macrophages, initially via surface-exposed pattern recognition receptors; activation of these cells increases following internalization and degradation of spirochetes within phagolysosomes. DCs that have taken up spirochetes migrate to the lymph nodes, where they present processed borrelial antigens to T cells and B cells. Sensitized T cells enter the circulation and are recruited to the site of infection. Plasma cells secrete specific antibodies that can kill B. burgdorferi via complement-dependent and –independent pathways. Production of pro-inflammatory cytokines by activated macrophages results in the recruitment of additional neutrophils, T cells, macrophages and DCs to the bite site, and eventually the development of erythema migrans. OspC, outer surface lipoprotein C; TLR, Toll-like receptor. .
References
-
- Benach JL, Garcia Monco JC. In: Borrelia: Molecular Biology, Host Interaction, and Pathogenesis. Samuels DS, Radolf JD, editors. Caister Academic; Norfolk, UK: 2010. pp. 7–26.
-
- Steere AC, et al. Lyme arthritis: an epidemic of oligoarticular arthritis in children and adults in three Connecticut communities. Arthritis Rheum. 1977;20:7–17. This article describes the original epidemiological study of the outbreak of arthritis in and around Lyme, Connecticut. - PubMed
-
- Burgdorfer W, et al. Lyme disease—a tick-borne spirochetosis? Science. 1982;216:1317–1319. This classic paper provides the first convincing evidence that Lyme disease is a tick-borne illness. - PubMed
-
- Steere AC, et al. The spirochetal etiology of Lyme disease. N Engl J Med. 1983;308:733–740. - PubMed
-
- Benach JL, et al. Spirochetes isolated from the blood of two patients with Lyme disease. N Engl J Med. 1983;308:740–742. This and reference 4 are the first two reports to describe the isolation of Lyme disease spirochaetes from patients. - PubMed
Publication types
MeSH terms
Grants and funding
- AI044254/AI/NIAID NIH HHS/United States
- R01 AI071107/AI/NIAID NIH HHS/United States
- R03 AI085248/AI/NIAID NIH HHS/United States
- R56 AI080646/AI/NIAID NIH HHS/United States
- AI068799/AI/NIAID NIH HHS/United States
- AI085248/AI/NIAID NIH HHS/United States
- AI080646/AI/NIAID NIH HHS/United States
- R56 AI044254/AI/NIAID NIH HHS/United States
- AI082436/AI/NIAID NIH HHS/United States
- R21 AI082436/AI/NIAID NIH HHS/United States
- AI029735/AI/NIAID NIH HHS/United States
- AI071107/AI/NIAID NIH HHS/United States
- R01 AI044254/AI/NIAID NIH HHS/United States
- R41 AI078631/AI/NIAID NIH HHS/United States
- R56 AI029735/AI/NIAID NIH HHS/United States
- R01 AI029735/AI/NIAID NIH HHS/United States
- AI029735‑20S1/AI/NIAID NIH HHS/United States
- R01 AI068799/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
