

Cysteinyl leukotriene type I receptor desensitization sustains Ca2+-dependent gene expression
- PMID: 22230957
- PMCID: PMC3272478
- DOI: 10.1038/nature10731
Cysteinyl leukotriene type I receptor desensitization sustains Ca2+-dependent gene expression
Abstract
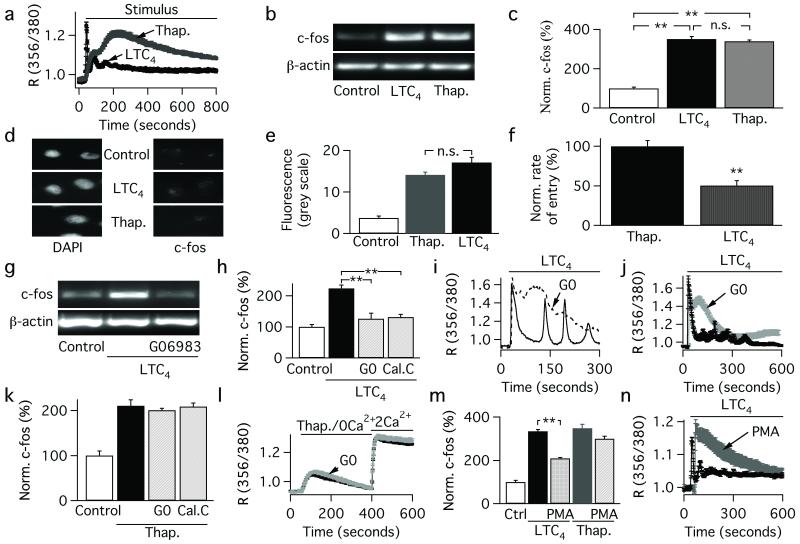
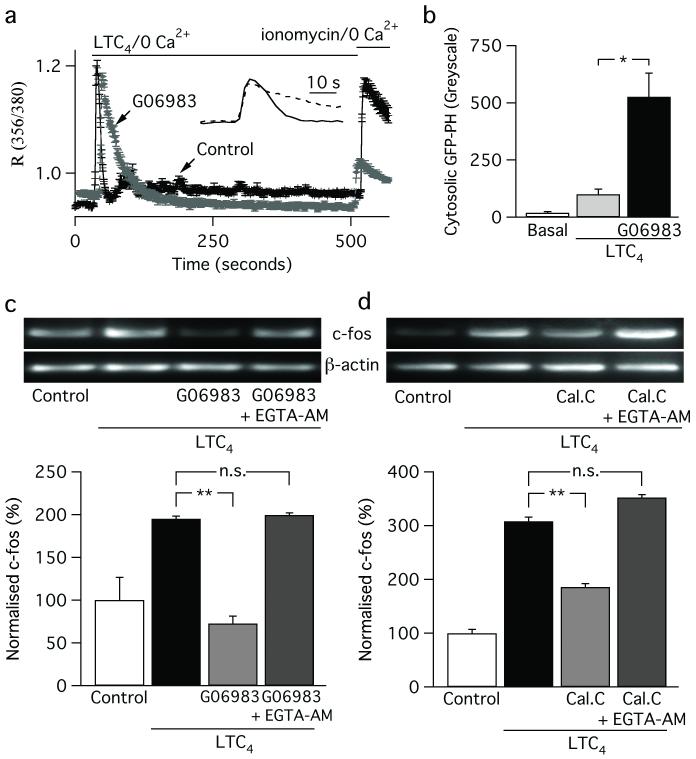
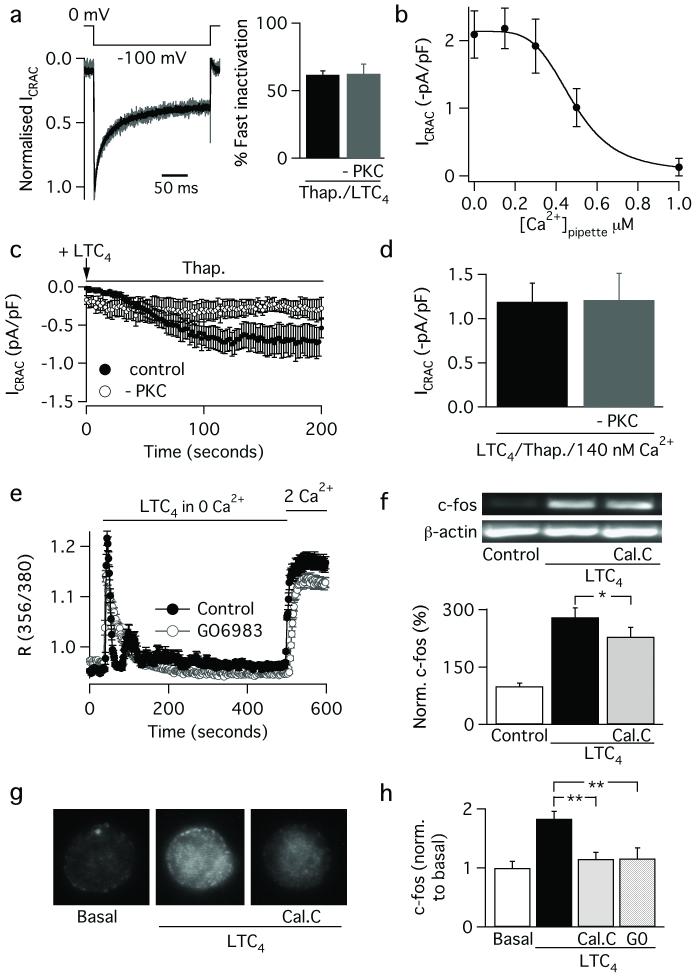
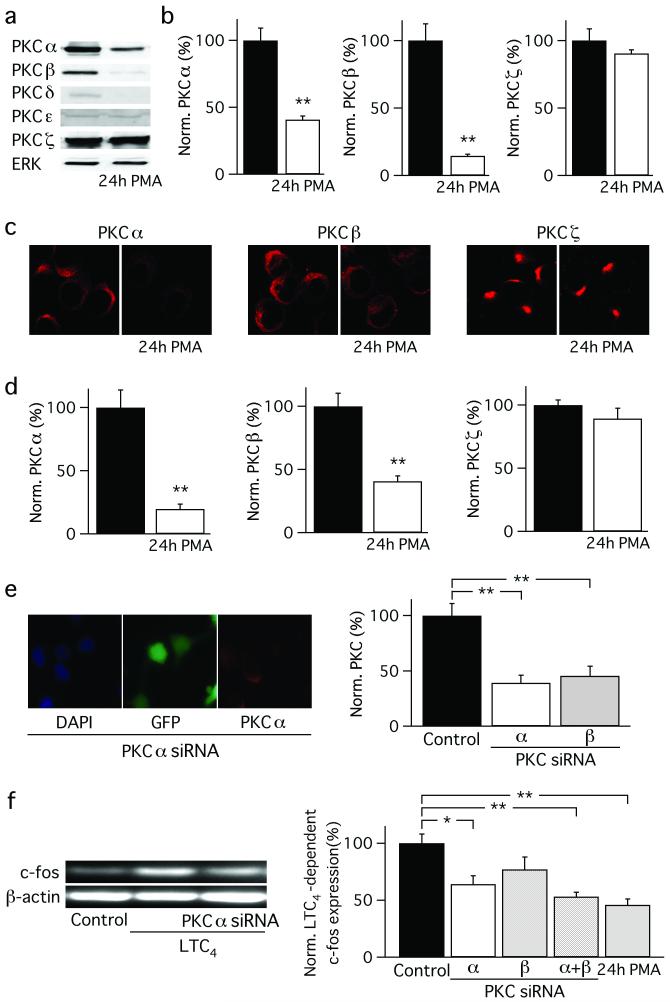
Receptor desensitization is a universal mechanism to turn off a biological response; in this process, the ability of a physiological trigger to activate a cell is lost despite the continued presence of the stimulus. Receptor desensitization of G-protein-coupled receptors involves uncoupling of the receptor from its G-protein or second-messenger pathway followed by receptor internalization. G-protein-coupled cysteinyl leukotriene type I (CysLT1) receptors regulate immune-cell function and CysLT1 receptors are an established therapeutic target for allergies, including asthma. Desensitization of CysLT1 receptors arises predominantly from protein-kinase-C-dependent phosphorylation of three serine residues in the receptor carboxy terminus. Physiological concentrations of the receptor agonist leukotriene C(4) (LTC(4)) evoke repetitive cytoplasmic Ca(2+) oscillations, reflecting regenerative Ca(2+) release from stores, which is sustained by Ca(2+) entry through store-operated calcium-release-activated calcium (CRAC) channels. CRAC channels are tightly linked to expression of the transcription factor c-fos, a regulator of numerous genes important to cell growth and development. Here we show that abolishing leukotriene receptor desensitization suppresses agonist-driven gene expression in a rat cell line. Mechanistically, stimulation of non-desensitizing receptors evoked prolonged inositol-trisphosphate-mediated Ca(2+) release, which led to accelerated Ca(2+)-dependent slow inactivation of CRAC channels and a subsequent loss of excitation-transcription coupling. Hence, rather than serving to turn off a biological response, reversible desensitization of a Ca(2+) mobilizing receptor acts as an 'on' switch, sustaining long-term signalling in the immune system.
Figures
Similar articles
-
Caveolin-1 alters the pattern of cytoplasmic Ca2+ oscillations and Ca2+-dependent gene expression by enhancing leukotriene receptor desensitization.J Biol Chem. 2014 Jun 20;289(25):17843-53. doi: 10.1074/jbc.M114.553453. Epub 2014 Apr 22. J Biol Chem. 2014. PMID: 24755228 Free PMC article.
-
Ca2+ Influx through Store-operated Calcium Channels Replenishes the Functional Phosphatidylinositol 4,5-Bisphosphate Pool Used by Cysteinyl Leukotriene Type I Receptors.J Biol Chem. 2015 Dec 4;290(49):29555-66. doi: 10.1074/jbc.M115.678292. Epub 2015 Oct 14. J Biol Chem. 2015. PMID: 26468289 Free PMC article.
-
Intercellular Ca2+ wave propagation involving positive feedback between CRAC channels and cysteinyl leukotrienes.FASEB J. 2009 Mar;23(3):894-905. doi: 10.1096/fj.08-118935. Epub 2008 Oct 31. FASEB J. 2009. PMID: 18978154
-
Store-Independent Orai Channels Regulated by STIM.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 11. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 11. PMID: 30299650 Free Books & Documents. Review.
-
Modulation of Orai1 and STIM1 by Cellular Factors.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 4. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 4. PMID: 30299655 Free Books & Documents. Review.
Cited by
-
Differential regulation of cysteinyl leukotriene receptor signaling by protein kinase C in human mast cells.PLoS One. 2013 Aug 15;8(8):e71536. doi: 10.1371/journal.pone.0071536. eCollection 2013. PLoS One. 2013. PMID: 23977066 Free PMC article.
-
Role of the Purinergic P2Y2 Receptor in Pulmonary Hypertension.Int J Environ Res Public Health. 2021 Oct 20;18(21):11009. doi: 10.3390/ijerph182111009. Int J Environ Res Public Health. 2021. PMID: 34769531 Free PMC article. Review.
-
The whole-cell Ca2+ release-activated Ca2+ current, ICRAC , is regulated by the mitochondrial Ca2+ uniporter channel and is independent of extracellular and cytosolic Na.J Physiol. 2020 May;598(9):1753-1773. doi: 10.1113/JP276551. Epub 2019 Feb 6. J Physiol. 2020. PMID: 30582626 Free PMC article.
-
The role of Orai-STIM calcium channels in melanocytes and melanoma.J Physiol. 2016 Jun 1;594(11):2825-35. doi: 10.1113/JP271141. Epub 2016 Apr 6. J Physiol. 2016. PMID: 26864956 Free PMC article. Review.
-
Ca(2+) Channel Re-localization to Plasma-Membrane Microdomains Strengthens Activation of Ca(2+)-Dependent Nuclear Gene Expression.Cell Rep. 2015 Jul 14;12(2):203-16. doi: 10.1016/j.celrep.2015.06.018. Epub 2015 Jul 2. Cell Rep. 2015. PMID: 26146085 Free PMC article.
References
-
- Ng S-W, DiCapite JL, Singaravelu K, Parekh AB. Sustained activation of the tyrosine kinase Syk by antigen in mast cells requires local Ca2+ influx through Ca2+ release-activated Ca2+ channels. Journal of Biological Chemistry. 2008;283:31348–31355. - PubMed
-
- DiCapite JL, Shirley A, Nelson C, Bates G, Parekh AB. Intercellular calcium wave propagation involving positive feedback between CRAC channels and cysteinyl leukotrienes. FASEB Journal. 2009;23:894–905. - PubMed
-
- Pierce KL, Premont RT, Lefkowitz RJ. Seven-transmembrane receptors. Nature Reviews Mol. Cell Biology. 2002;3:639–650. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
