

Timing facilitated site transfer of an enzyme on DNA
- PMID: 22231272
- PMCID: PMC3262087
- DOI: 10.1038/nchembio.764
Timing facilitated site transfer of an enzyme on DNA
Abstract
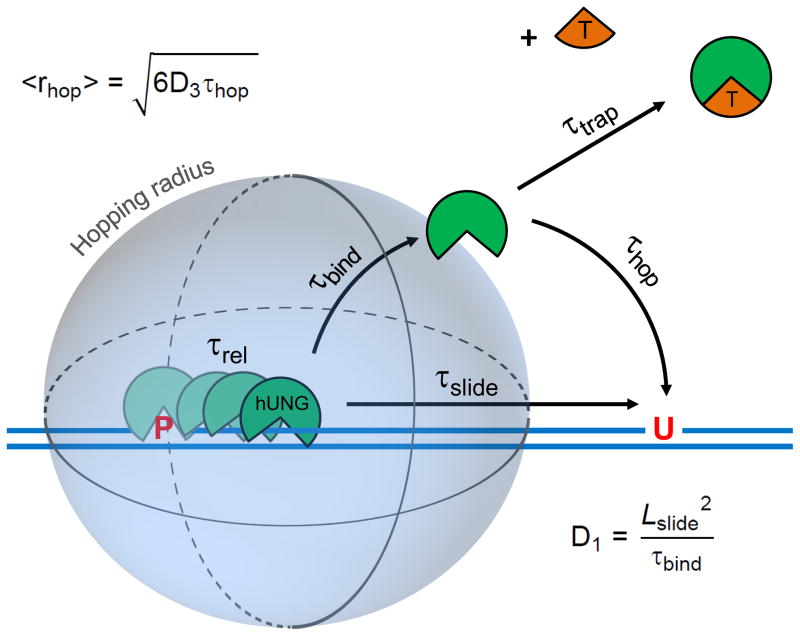
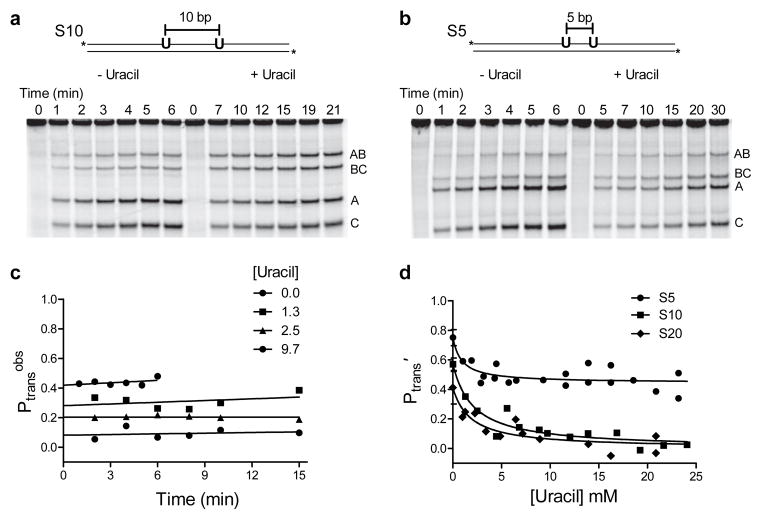
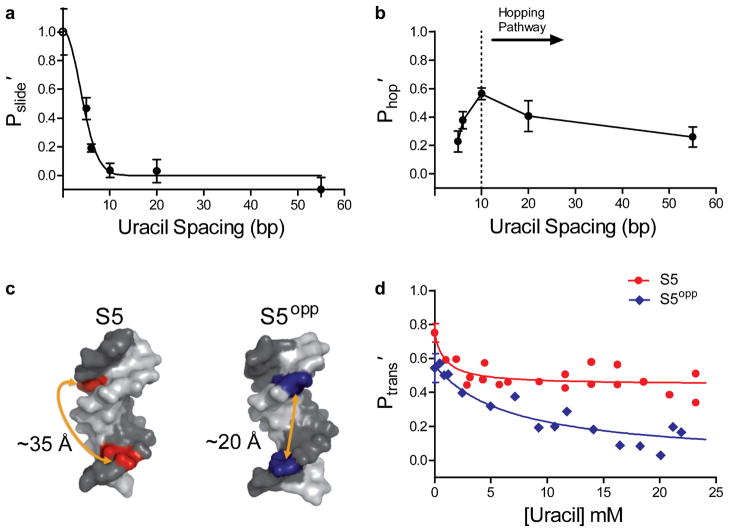
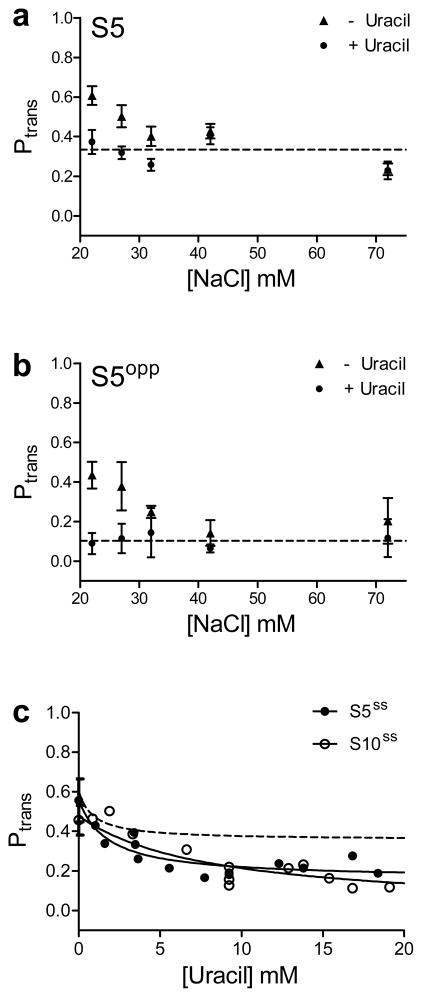
Many enzymes that react with specific sites in DNA have the property of facilitated diffusion, in which the DNA chain is used as a conduit to accelerate site location. Despite the importance of such mechanisms in gene regulation and DNA repair, there have been few viable approaches to elucidate the microscopic process of facilitated diffusion. Here we describe a new method in which a small-molecule trap (uracil) is used to clock a DNA repair enzyme as it hops and slides between damaged sites in DNA. The 'molecular clock' provides unprecedented information: the mean length for DNA sliding, the one-dimensional diffusion constant, the maximum hopping radius and the time frame for DNA hopping events. In addition, the data establish that the DNA phosphate backbone is a sufficient requirement for DNA sliding.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
