

Chk1 phosphorylation of Metnase enhances DNA repair but inhibits replication fork restart
- PMID: 22231448
- PMCID: PMC3963179
- DOI: 10.1038/onc.2011.586
Chk1 phosphorylation of Metnase enhances DNA repair but inhibits replication fork restart
Erratum in
- Oncogene. 2014 Jan 23;33(4):536. Laitao, A [corrected to Leitao, A]
Abstract
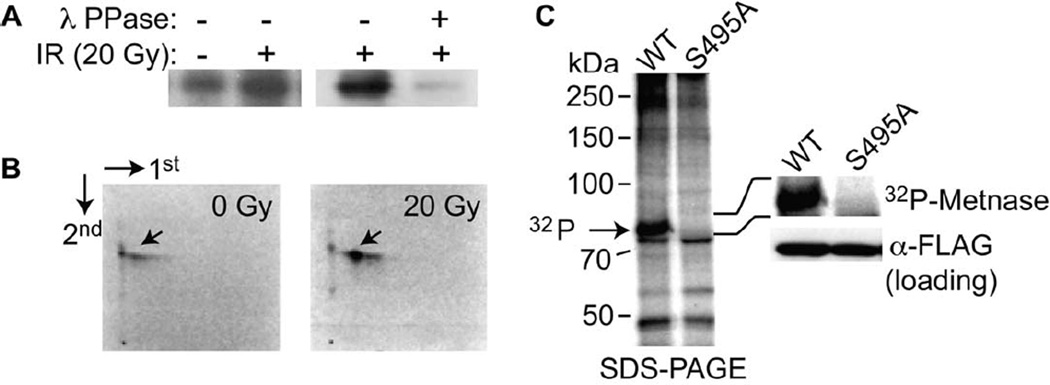
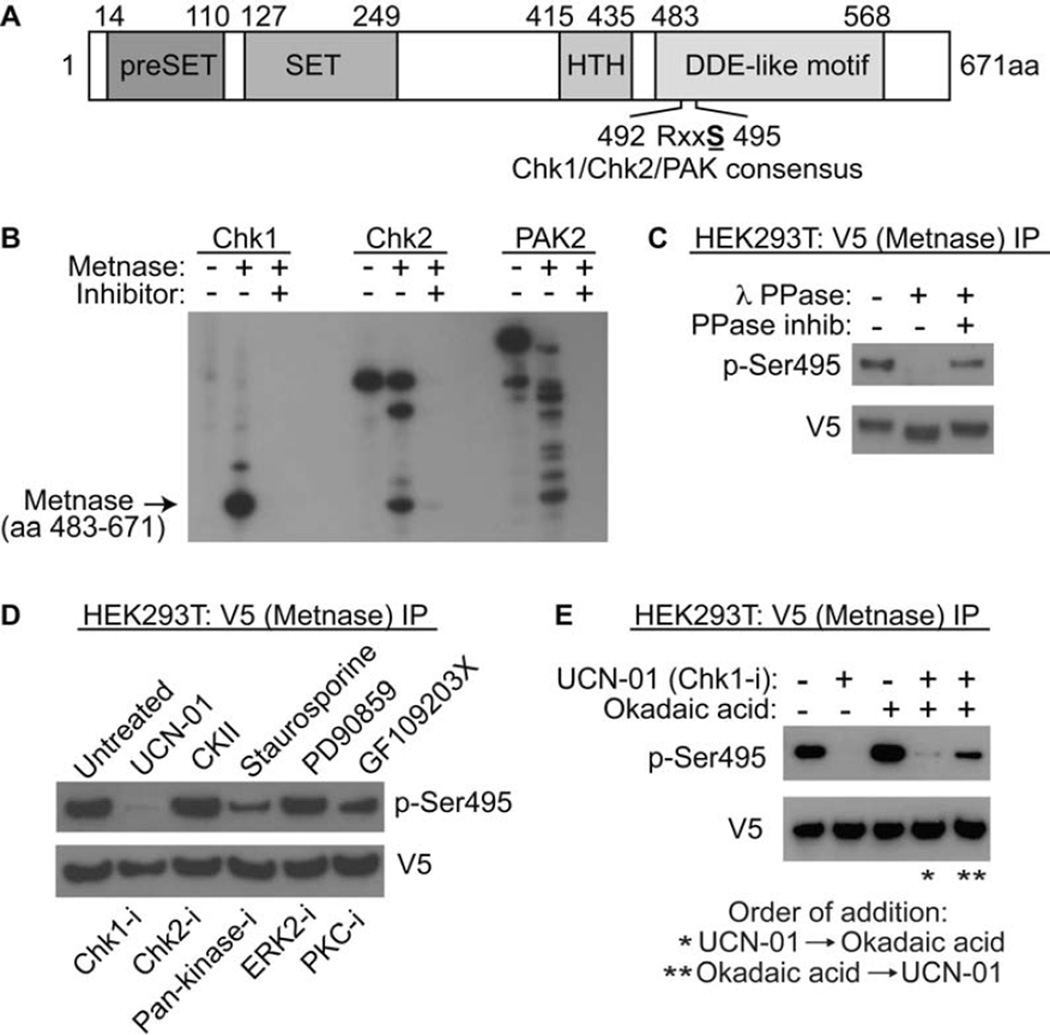
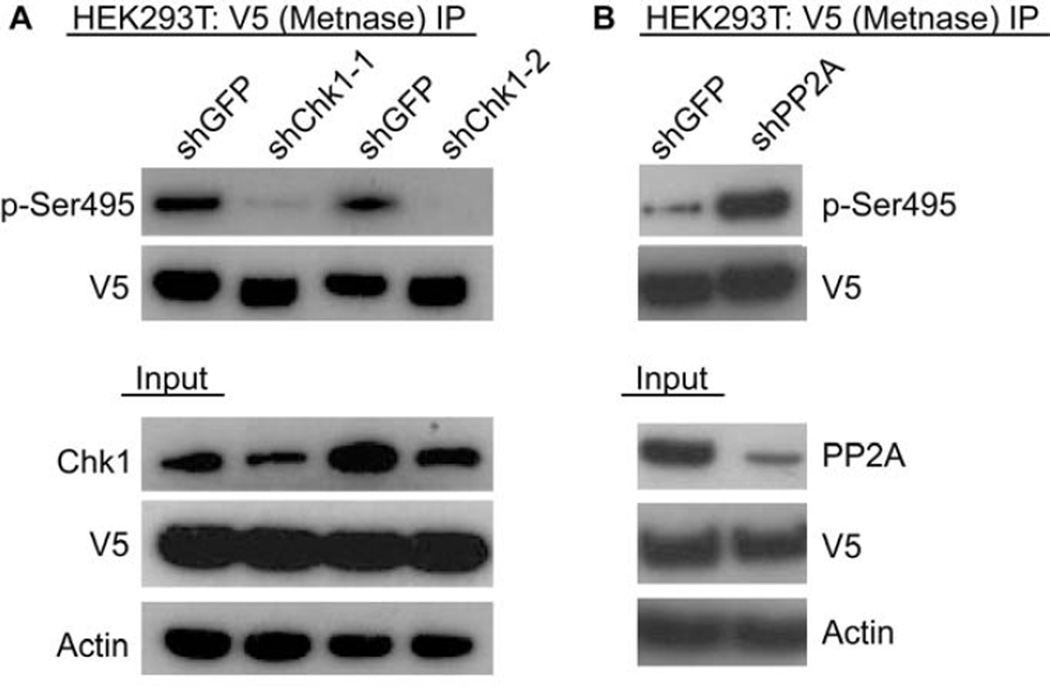
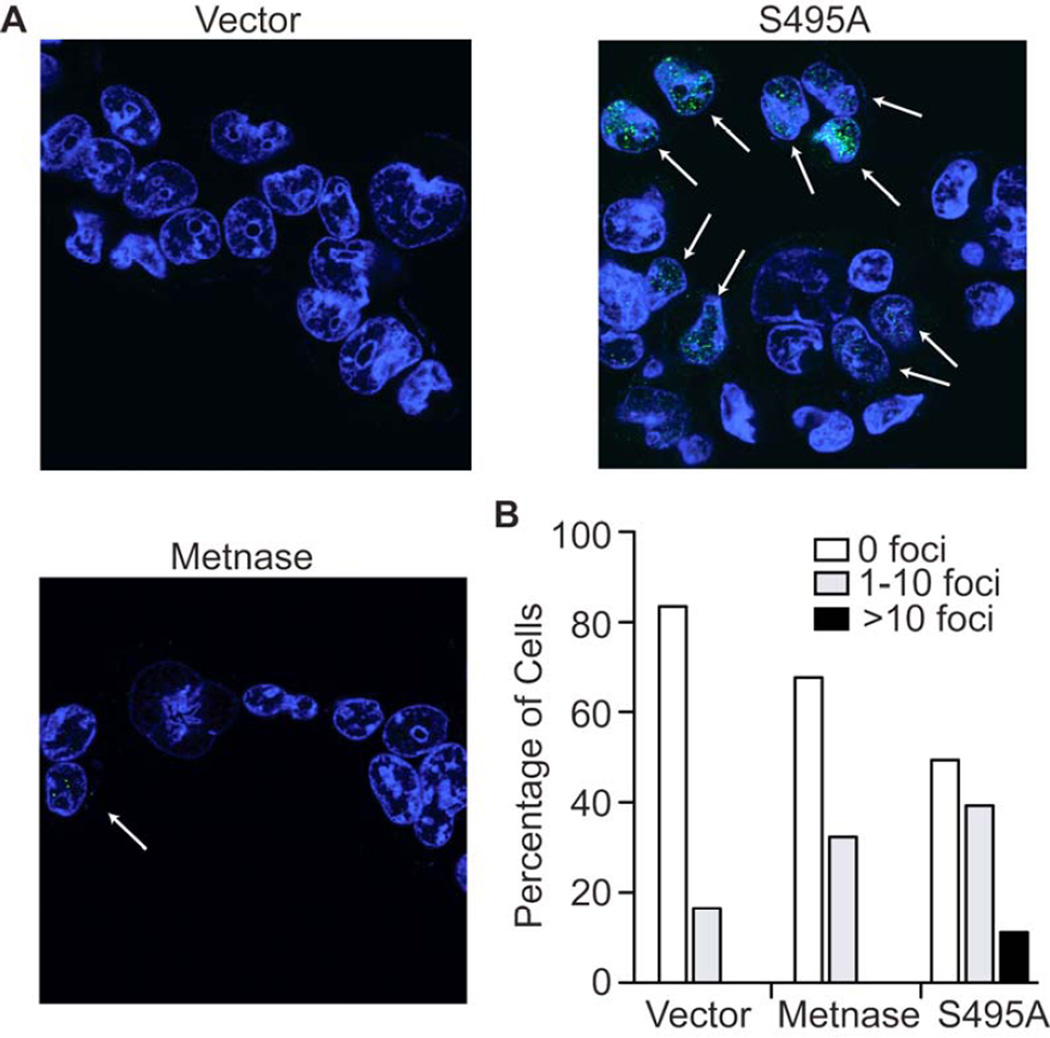
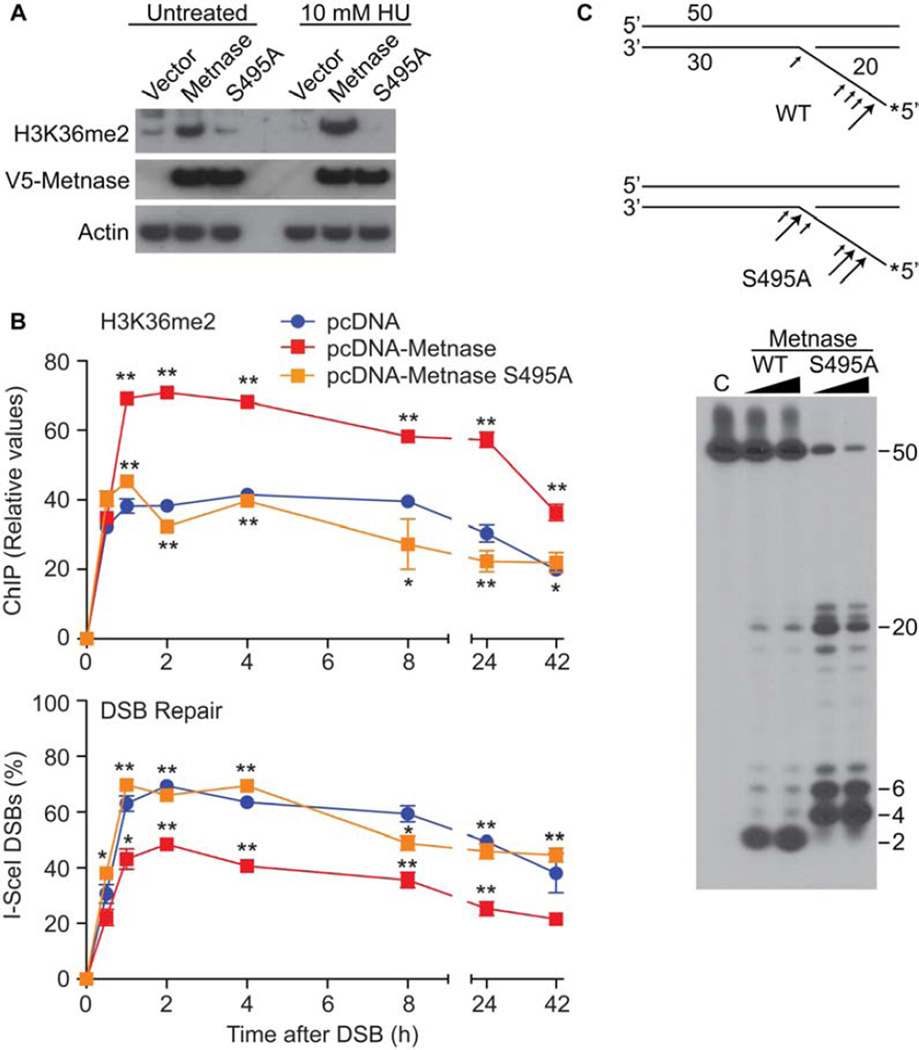
Chk1 both arrests replication forks and enhances repair of DNA damage by phosphorylating downstream effectors. Although there has been a concerted effort to identify effectors of Chk1 activity, underlying mechanisms of effector action are still being identified. Metnase (also called SETMAR) is a SET and transposase domain protein that promotes both DNA double-strand break (DSB) repair and restart of stalled replication forks. In this study, we show that Metnase is phosphorylated only on Ser495 (S495) in vivo in response to DNA damage by ionizing radiation. Chk1 is the major mediator of this phosphorylation event. We had previously shown that wild-type (wt) Metnase associates with chromatin near DSBs and methylates histone H3 Lys36. Here we show that a Ser495Ala (S495A) Metnase mutant, which is not phosphorylated by Chk1, is defective in DSB-induced chromatin association. The S495A mutant also fails to enhance repair of an induced DSB when compared with wt Metnase. Interestingly, the S495A mutant demonstrated increased restart of stalled replication forks compared with wt Metnase. Thus, phosphorylation of Metnase S495 differentiates between these two functions, enhancing DSB repair and repressing replication fork restart. In summary, these data lend insight into the mechanism by which Chk1 enhances repair of DNA damage while at the same time repressing stalled replication fork restart.
Figures

Similar articles
-
The DNA repair component Metnase regulates Chk1 stability.Cell Div. 2014 Jul 9;9:1. doi: 10.1186/1747-1028-9-1. eCollection 2014. Cell Div. 2014. PMID: 25024738 Free PMC article.
-
Metnase Mediates Loading of Exonuclease 1 onto Single Strand Overhang DNA for End Resection at Stalled Replication Forks.J Biol Chem. 2017 Jan 27;292(4):1414-1425. doi: 10.1074/jbc.M116.745646. Epub 2016 Dec 14. J Biol Chem. 2017. PMID: 27974460 Free PMC article.
-
Metnase/SETMAR: a domesticated primate transposase that enhances DNA repair, replication, and decatenation.Genetica. 2010 May;138(5):559-66. doi: 10.1007/s10709-010-9452-1. Epub 2010 Mar 23. Genetica. 2010. PMID: 20309721 Free PMC article. Review.
-
Metnase promotes restart and repair of stalled and collapsed replication forks.Nucleic Acids Res. 2010 Sep;38(17):5681-91. doi: 10.1093/nar/gkq339. Epub 2010 May 10. Nucleic Acids Res. 2010. PMID: 20457750 Free PMC article.
-
The fork and the kinase: a DNA replication tale from a CHK1 perspective.Mutat Res Rev Mutat Res. 2015 Jan-Mar;763:168-80. doi: 10.1016/j.mrrev.2014.10.003. Epub 2014 Oct 22. Mutat Res Rev Mutat Res. 2015. PMID: 25795119 Free PMC article. Review.
Cited by
-
cAMP protein kinase phosphorylates the Mos1 transposase and regulates its activity: evidences from mass spectrometry and biochemical analyses.Nucleic Acids Res. 2014 Jan;42(2):1117-28. doi: 10.1093/nar/gkt874. Epub 2013 Sep 29. Nucleic Acids Res. 2014. PMID: 24081583 Free PMC article.
-
SETMAR, a case of primate co-opted genes: towards new perspectives.Mob DNA. 2022 Apr 8;13(1):9. doi: 10.1186/s13100-022-00267-1. Mob DNA. 2022. PMID: 35395947 Free PMC article. Review.
-
The DDN catalytic motif is required for Metnase functions in non-homologous end joining (NHEJ) repair and replication restart.J Biol Chem. 2014 Apr 11;289(15):10930-10938. doi: 10.1074/jbc.M113.533216. Epub 2014 Feb 25. J Biol Chem. 2014. PMID: 24573677 Free PMC article.
-
Potential role for the Metnase transposase fusion gene in colon cancer through the regulation of key genes.PLoS One. 2014 Oct 15;9(10):e109741. doi: 10.1371/journal.pone.0109741. eCollection 2014. PLoS One. 2014. PMID: 25333365 Free PMC article.
-
Epigenetics in radiation-induced fibrosis.Oncogene. 2015 Apr 23;34(17):2145-55. doi: 10.1038/onc.2014.145. Epub 2014 Jun 9. Oncogene. 2015. PMID: 24909163 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
