

Role of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter baumannii strain ATCC 19606T with human lung epithelial cells, Galleria mellonella caterpillars, and mice
- PMID: 22232188
- PMCID: PMC3294665
- DOI: 10.1128/IAI.06279-11
Role of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter baumannii strain ATCC 19606T with human lung epithelial cells, Galleria mellonella caterpillars, and mice
Abstract
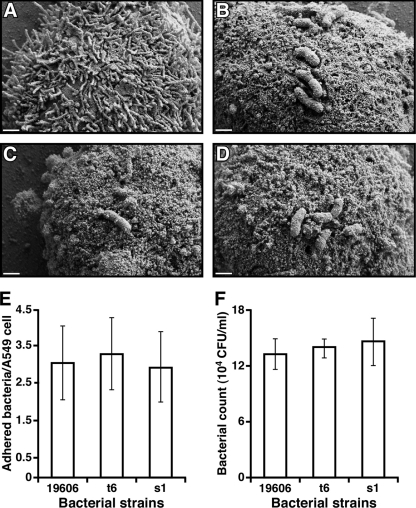
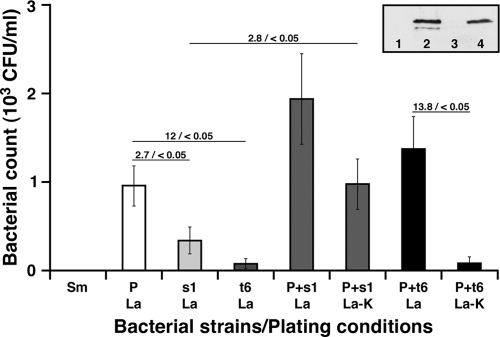
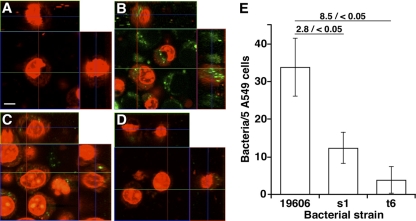
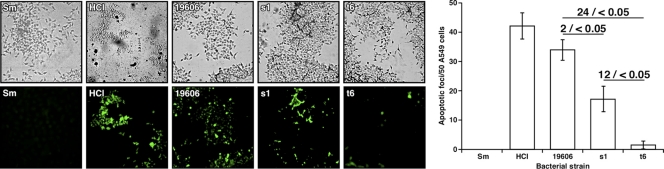
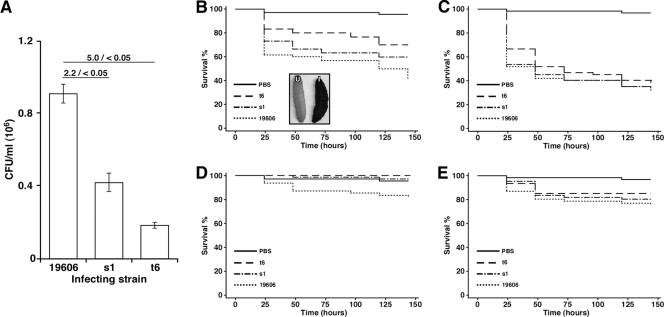
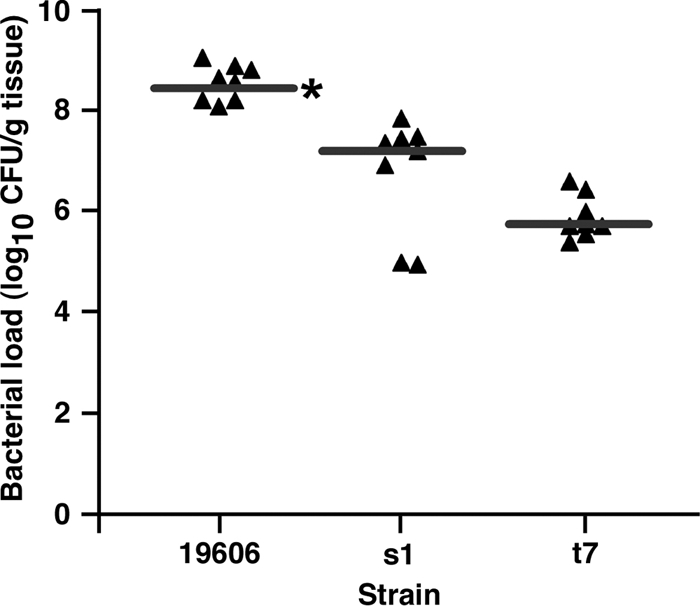
Acinetobacter baumannii, which causes serious infections in immunocompromised patients, expresses high-affinity iron acquisition functions needed for growth under iron-limiting laboratory conditions. In this study, we determined that the initial interaction of the ATCC 19606(T) type strain with A549 human alveolar epithelial cells is independent of the production of BasD and BauA, proteins needed for acinetobactin biosynthesis and transport, respectively. In contrast, these proteins are required for this strain to persist within epithelial cells and cause their apoptotic death. Infection assays using Galleria mellonella larvae showed that impairment of acinetobactin biosynthesis and transport functions significantly reduces the ability of ATCC 19606(T) cells to persist and kill this host, a defect that was corrected by adding inorganic iron to the inocula. The results obtained with these ex vivo and in vivo approaches were validated using a mouse sepsis model, which showed that expression of the acinetobactin-mediated iron acquisition system is critical for ATCC 19606(T) to establish an infection and kill this vertebrate host. These observations demonstrate that the virulence of the ATCC 19606(T) strain depends on the expression of a fully active acinetobactin-mediated system. Interestingly, the three models also showed that impairment of BasD production results in an intermediate virulence phenotype compared to those of the parental strain and the BauA mutant. This observation suggests that acinetobactin intermediates or precursors play a virulence role, although their contribution to iron acquisition is less relevant than that of mature acinetobactin.
Figures
References
-
- Abbott A. 2005. Medics braced for fresh superbug. Nature 436:758. - PubMed
-
- Antunes LC, Imperi F, Towner KJ, Visca P. 2011. Genome-assisted identification of putative iron-utilization genes in Acinetobacter baumannii and their distribution among a genotypically diverse collection of clinical isolates. Res. Microbiol. 162:279–284 - PubMed
-
- Brachelente C, Wiener D, Malik Y, Huessy D. 2007. A case of necrotizing fasciitis with septic shock in a cat caused by Acinetobacter baumannii. Vet. Dermatol. 18:432–438 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
