

Hemin binding protein C is found in outer membrane vesicles and protects Bartonella henselae against toxic concentrations of hemin
- PMID: 22232189
- PMCID: PMC3294634
- DOI: 10.1128/IAI.05769-11
Hemin binding protein C is found in outer membrane vesicles and protects Bartonella henselae against toxic concentrations of hemin
Abstract
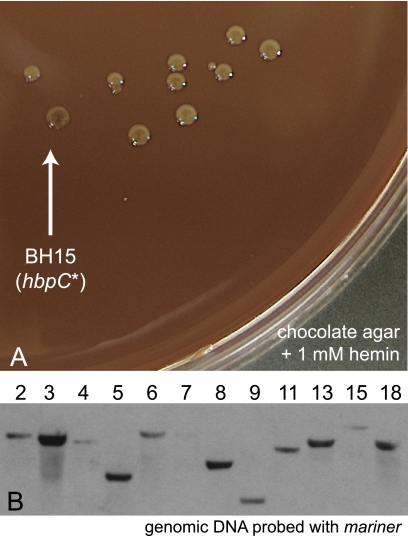
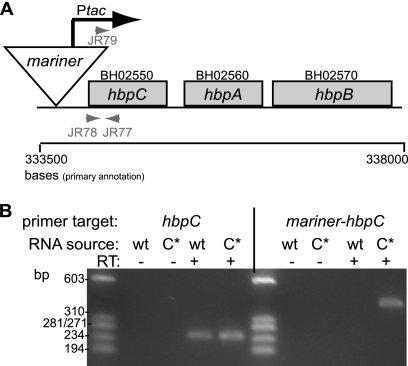
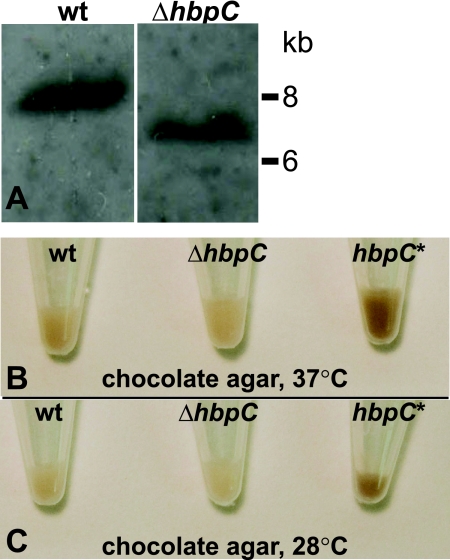
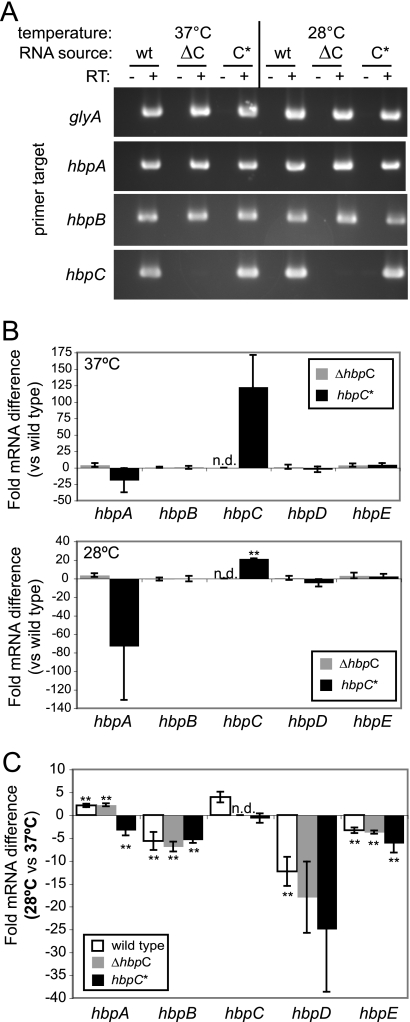
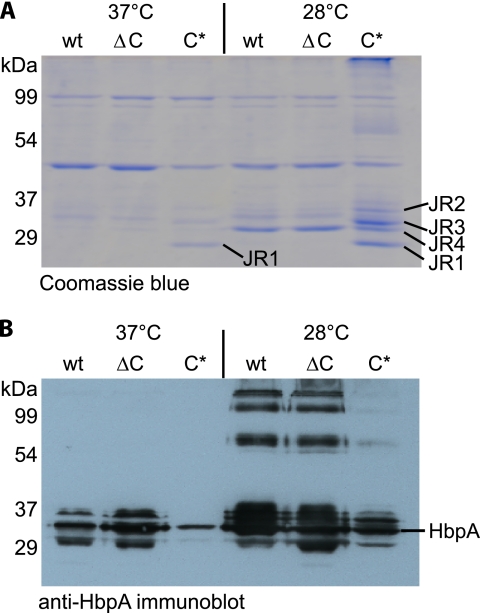
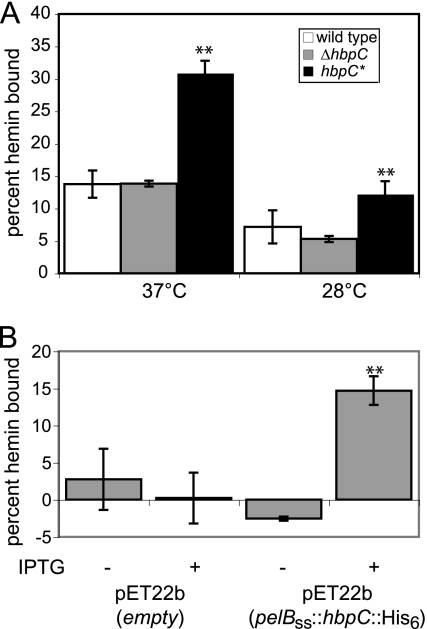
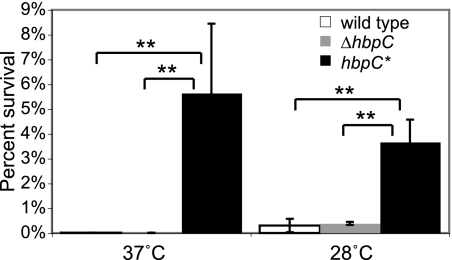
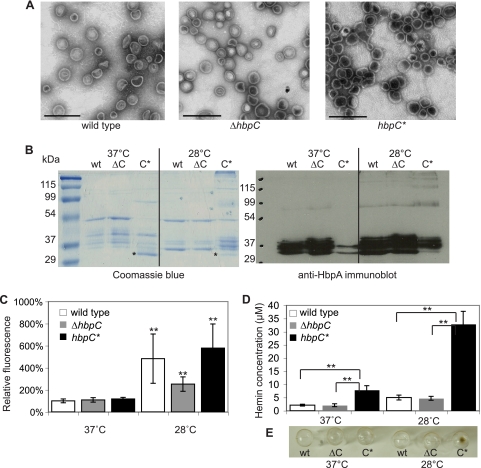
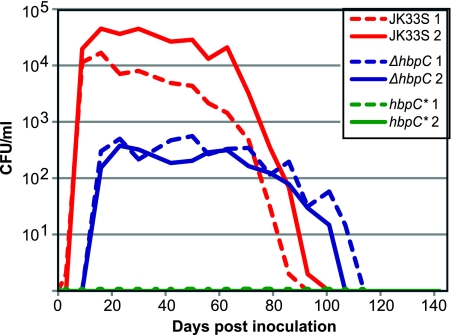
Bartonella species are gram-negative, emerging bacterial pathogens found in two distinct environments. In the gut of the obligately hematophagous arthropod vector, bartonellae are exposed to concentrations of heme that are toxic to other bacteria. In the bloodstream of the mammalian host, access to heme and iron is severely restricted. Bartonellae have unusually high requirements for heme, which is their only utilizable source of iron. Although heme is essential for Bartonella survival, little is known about genes involved in heme acquisition and detoxification. We developed a strategy for high-efficiency transposon mutagenesis to screen for genes in B. henselae heme binding and uptake pathways. We identified a B. henselae transposon mutant that constitutively expresses the hemin binding protein C (hbpC) gene. In the wild-type strain, transcription of B. henselae hbpC was upregulated at arthropod temperature (28°C), compared to mammalian temperature (37°C). In the mutant strain, temperature-dependent regulation was absent. We demonstrated that HbpC binds hemin and localizes to the B. henselae outer membrane and outer membrane vesicles. Overexpression of hbpC in B. henselae increased resistance to heme toxicity, implicating HbpC in protection of B. henselae from the toxic levels of heme present in the gut of the arthropod vector. Experimental inoculation of cats with B. henselae strains demonstrated that both constitutive expression and deletion of hbpC affect the ability of B. henselae to infect the cat host. Modulation of hbpC expression appears to be a strategy employed by B. henselae to survive in the arthropod vector and the mammalian host.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
